Maxell
-
Postów
46 214 -
Dołączył
-
Ostatnia wizyta
-
Wygrane w rankingu
180
Typ zawartości
Profile
Forum
Wydarzenia
Mapa użytkowników
Sklep
Giełda
Treść opublikowana przez Maxell
-
[Lektura obowiązkowa] Wędzenie wyrobów
Maxell odpowiedział(a) na Maxell temat w Wędzarnie. Budowa i obsługa
Andyandy, chyba nie masz zamiaru polemizować z prof. Pezackim? To byłaby już lekka przesada. Poza tym, wszystko na temat wędzenia i wędzarni zostało już dawno wymyślone. Wkleiłem (i mam nadzieję dalej to robić) materiały, które wyczerpują temat technologii wędzenia, budowy wędzarni, sposobów produkcji wędzonek itp. Wybieram materiały opracowane przez ludzi, którzy całe swe życie poświęcili badaniu tego zagadnienia i mieli zawsze oparcie w najlepszych laboratoriach. Wyniki badań są podstawą do oceniania i wydawania ostatecznych opinii. To podstawa. Bez uczciwych badań jedynie sobie gdybamy, a dyskusja jest czysto akademicka. -
[Lektura obowiązkowa] Wędzenie wyrobów
Maxell odpowiedział(a) na Maxell temat w Wędzarnie. Budowa i obsługa
Jeśli wędzisz na zimno, to tak. Po to została zbudowana. -
[Lektura obowiązkowa] Wędzenie wyrobów
Maxell odpowiedział(a) na Maxell temat w Wędzarnie. Budowa i obsługa
Zasady Kolego zasady. -
[Lektura obowiązkowa] Wędzenie wyrobów
Maxell odpowiedział(a) na Maxell temat w Wędzarnie. Budowa i obsługa
Jakim normom? Każdy rodzaj wędzarni ma swoje plusy i minusy. Nie ma ideału. -
Musisz w takim razie zmienić forum.
-
[Lektura obowiązkowa] Wędzenie wyrobów
Maxell odpowiedział(a) na Maxell temat w Wędzarnie. Budowa i obsługa
A co tutaj jest skomplikowanego? Normalnie przedstawione jedne z podstawowych procesów technologicznych przy produkcji wyrobów masarskich wędzonych. Grillowaniu poświęcone są inne działy na forum. -
Wędzarnia z szafki, jaka wielkość paleniska i rury ?
Maxell odpowiedział(a) na marula3 temat w Wędzarnie. Budowa i obsługa
Zrób sobie pod szafą palenisko skrzyniowe (ruchome) takie, jakie zaprezentował w swojej składanej wędzarni Kazik55. -
Przenośna wędzarnia Kazika55 z paleniskiem skrzyniowym
Maxell opublikował(a) temat w Wędzarnie. Budowa i obsługa
Wszyscy uważają, że czegoś nie da się zrobić. Aż przychodzi taki jeden, który nie wie, że się nie da. I on właśnie to robi.



-
Bardzo ciekawe i warte wypróbowania
Maxell odpowiedział(a) na Maxell temat w Polska tradycja masarska
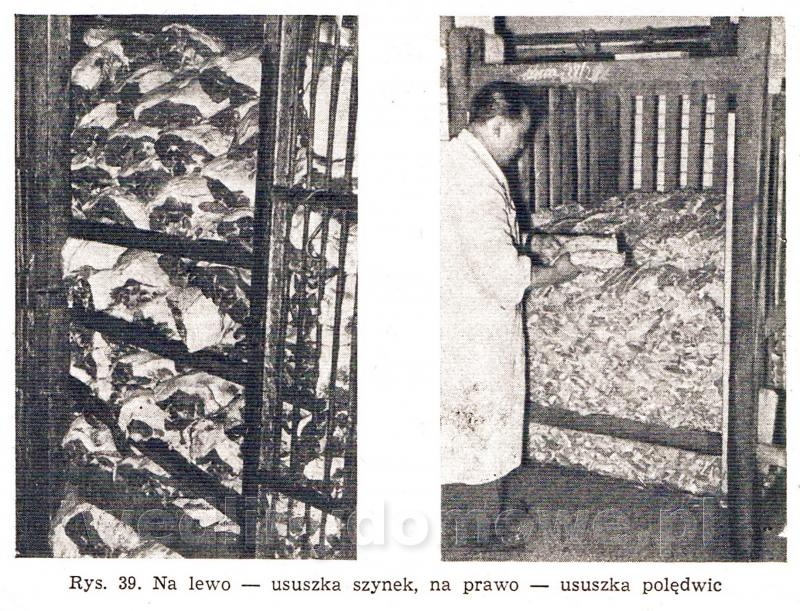
Do informacji z pierwszego posta, dodam dwa zdjęcia: 1. Kontrolowana ususzka szynek i polędwic: 2. Mycie szynek po okresie dojrzewania: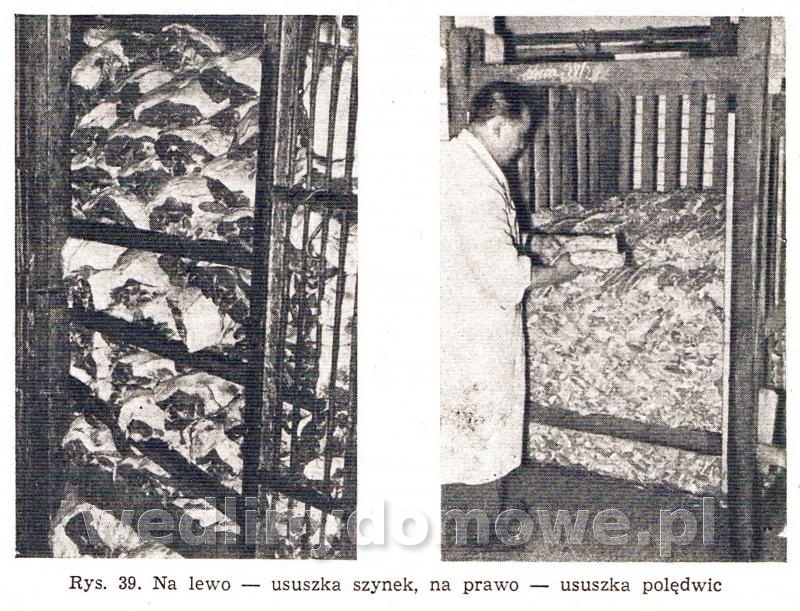

-
Dziękuję za sprostowanie. Podałem ten typ gołębników, gdyż z racji ukierunkowania naszego forum, interesują nas głównie rasy mięsne.
-
Sposób wędzenia mojego dziadka. Czy dobry?
Maxell odpowiedział(a) na karuzo temat w Dla początkujących
Jeśli masz kłopot z wrzucaniem zdjęć - podsyłasz je do mnie na pocztę i po problemie. -
O S U S Z A N I E !!! i Ewentualnie mokre paliwo.
-
Sposób wędzenia mojego dziadka. Czy dobry?
Maxell odpowiedział(a) na karuzo temat w Dla początkujących
Nieźle pobalowaliście przy tej robocie. -
http://sprzedajemy.pl/lodka-drewniana-nr21839062
-
Bardzo ciekawe i warte wypróbowania
Maxell odpowiedział(a) na Maxell temat w Polska tradycja masarska
No to mamy już wyjaśnioną zagadkę. -
Przygotowując materiały do wrzucenia na forum, natrafiłem na bardzo ciekawą książkę dr B. Godlewicza, pt.: "Dozór sanitarno-weterynaryjny nad mięsem i jego przetworami" (PWRiL, Warszawa 1959 r.). Jest tam m.in. potężny dział poświęcony peklowaniu mięsa. Tutaj właśnie natrafiłem na bardzo ciekawe informacje, m.in. dotycząca prowadzenia procesu tzw. ususzki (osuszki) mięsa po ukończonym peklowaniu mokrym. Proces ma za zadanie przede wszystkim, usunięcie wilgoci z mięsa. Prowadzi się go przez okres od kilku godzin do kilku tygodni w warunkach chłodnych (ale nie w chłodni) w środowisku dobrze pochłaniającym wilgoć. Poza tym chodzi w nim o wyrównanie zasolenia i barwy mięsa, a głównie zmianę smaku i zapachu tzw. dojrzewanie mięsa. Po ukończeniu dojrzewania moczy się mięso w zimnej lub ciepłej wodzie oraz zmywa tłuszcz i zanieczyszczenia skóry, co ułatwia wędzenie. Mięsa nie wolno myć szczotkami, gdyż może to spowodować jego zakażenie. Po takim umyciu mięso poddaje się osuszaniu w przewiewnych i chłodnych wędzarniach lub suszarniach. Mięso przed wędzeniem posypuje się niekiedy drobno zmielona solą lub mączka grochową. W Anglii i USA mięso, a specjalnie szynki, po osuszeniu wkłada się w woreczki ze specjalnej tkaniny (stockinetty) w celu zabezpieczenia go przed zanieczyszczeniami i owadami. Oczywiście mowa tutaj o mięsie peklowanym w solankach o stężeniu ok. 20 st. Be. Książka zawiera całą masę bardzo ciekawych informacji i najprawdopodobniej w całości zamieszczę ja na forum. A przy okazji wyjaśniła się tajemnica sposobu dojrzewania mięsa na steki..
-
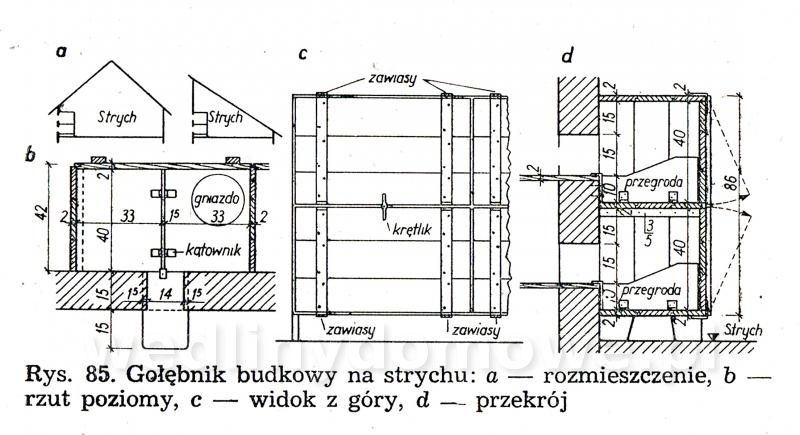
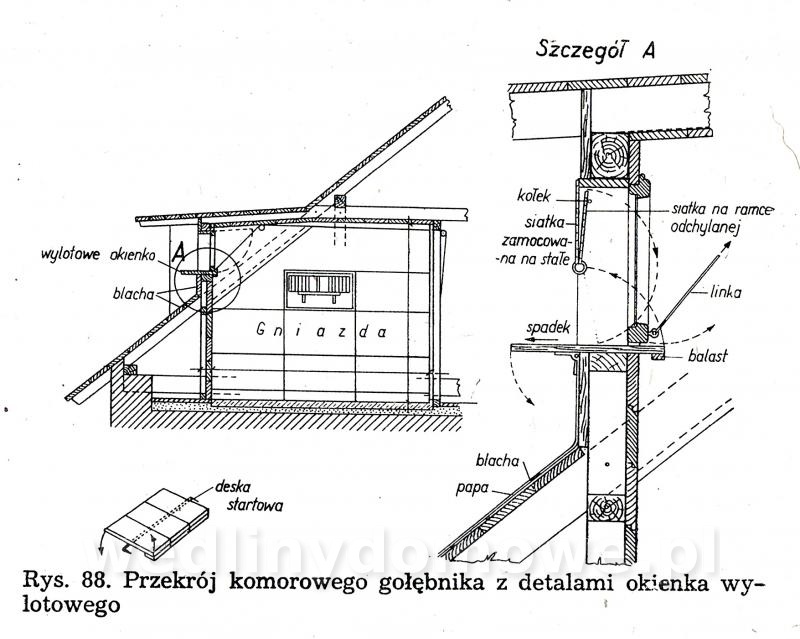
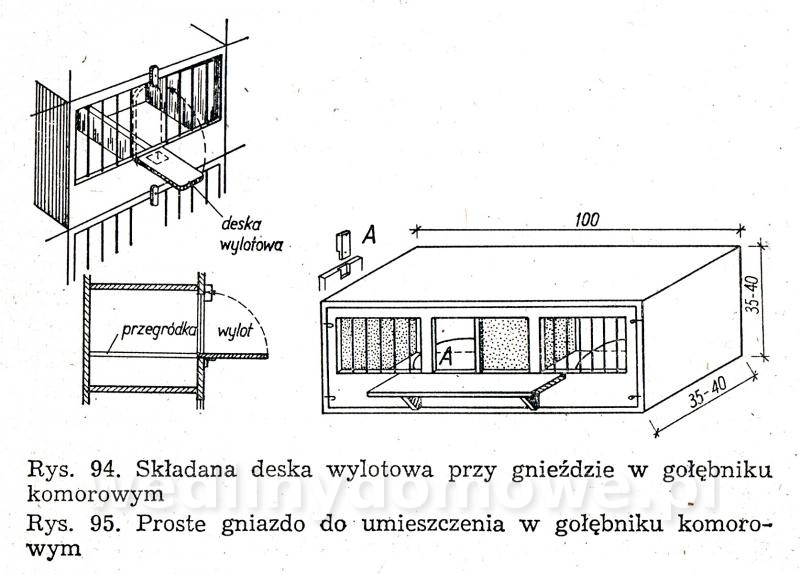
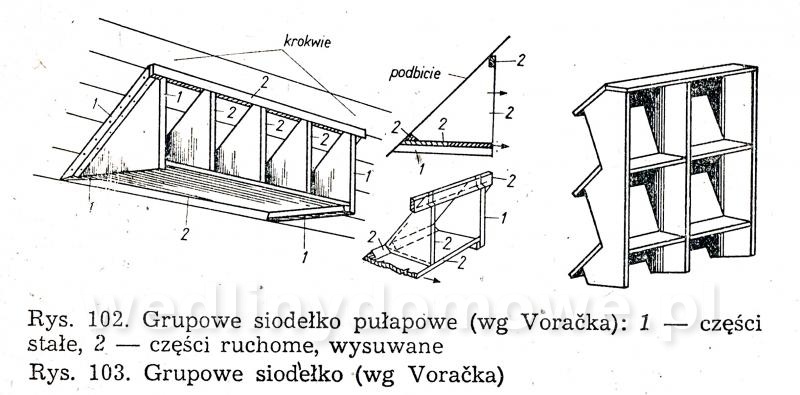
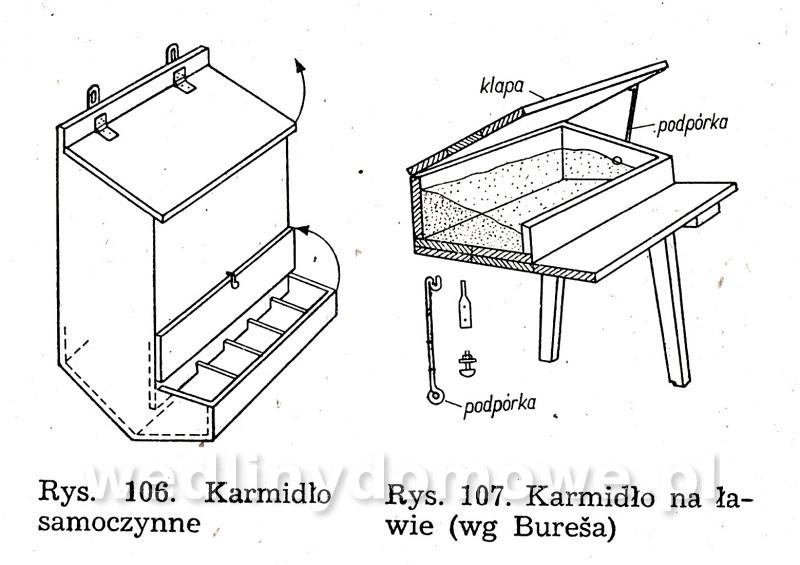
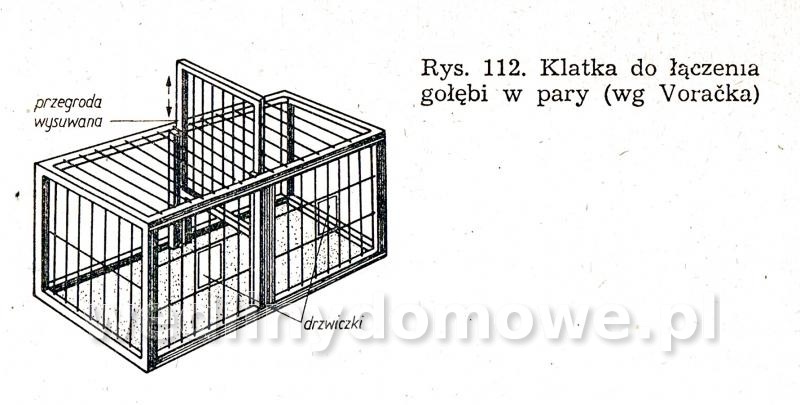
Gołębniki Bank zdjęć WB Doświadczony hodowca wie, że warunkiem osiągnięcia dobrych rezultatów hodowlanych jest zapewnienie gołębiom odpowiedniego pomieszczenia. Gołębnik powinien być dostatecznie duży, dobrze przewietrzany, ale nie powinno być w nim przeciągów, dobrze oświetlony, suchy i czysty. W lecie nie może się zbytnio nagrzewać, powinien być dla gołębi schronieniem przed nadmiernym słońcem. W gołębniku nie może być też zbyt ostre światło, a gniazda powinny być nawet w półmroku. Dlatego jeżeli tylko to możliwe, silne promienie słoneczne nie powinny docierać do gołębnika. Najlepiej, jeżeli przednia ściana jest zwrócona na wschód lub południowy wschód. Nie powinno się w ogóle budować gołębników zwróconych przednią ścianą na północ. Wylot z gołębnika powinien być widoczny z daleka, niczym nie osłonięty. Nie może być również zwrócony w stronę, z której najczęściej wieją wiatry. Dobrze również, jeżeli okienko wylotowe będzie widoczne z okien, mieszkania hodowcy. Może on wtedy częściej obserwować zachowanie się gołębi. Najczęściej sytuuje się gołębniki na strychach, tzw. gołębniki komorowe, lub buduje specjalnie w tym celu oddzielne komórki w ogródkach i na podwórkach. Można także budować gołębniki budkowe, umocowane na zewnętrznych ścianach budynków gospodarczych. Są one jednak mniej dogodne pod względem hodowlanym niż poprzednie. Wielkość gołębnika zależy od tego, jakiej rasy gołębie zamierzamy hodować. Długość poszczególnej komórki wynosi od 70 do 100 cm, szerokość od 35 do 40 cm, a wysokość od 35 do 40 cm. Dla dużych gołębi szerokość i wysokość komórki powinna wynosić około 50 cm. Gołębie muszą mieć pod dostatkiem czystego powietrza, dlatego gołębniki nie powinny być umieszczane w miejscach źle wietrzonych. Gołębniki budkowe. Umocowuje się je na zewnętrznych ścianach budynków, najczęściej nad okapem (rys. 82 i 83). Ściany silnie nasłonecznione, narażone na silne wiatry, a także ściany północne nie są odpowiednim miejscem do usytuowania gołębnika. Mogą być użytkowane tylko do chowu gołębi ras bardziej odpornych. Obecnie wykonuje się je tylko wtedy, kiedy miejscowe warunki (a niekiedy i finansowe) nie pozwalają na wykonanie innego gołębnika. Gołębniki takie trudno jest czyścić (z drabinki) i nie łatwo kontroluje się w nich gołębie. Trudności sprawia też chwytanie w nich gołębi (podczas przestawiania drabinki gołębie odlatują i często niechętnie wracają do gołębnika). Małe gołębie często wypadają z takich gołębników. Gołębnik budkowy powinien być umieszczony na suchej ścianie. Należy również osłonić go przed deszczem. Jeśli nie ma możliwości umieszczenia gołębnika na suchej ścianie, należy między jego tylną ścianę a mur włożyć izolacyjną wkładkę z papy. Powinna ona wystawać poza krawędź gołębnika na 3—5 cm. Gołębnik odsuwa się od ściany przez przymocowanie do jego tylnej ściany drewnianej listwy. Gołębniki zawieszane na ścianach chronimy przed deszczem daszkiem pokrytym papą. Jeśli gołębniki podokapowe umieszczone są pod dachem o nieszczelnym pokryciu dobrze jest również pokryć je papą. Gołębniki należy zabezpieczyć przed szkodnikami (koty, tchórze, kuny, łasiczki itd.) umieszczając je w miejscach trudno dostępnych dla szkodników, np. na gładkich ścianach, wyposażając je w znacznie wystający dach, umieszczając drzwiczki albo klapy w otworach wylotowych. Wylot można także zabezpieczyć umieszczając w nim obrotową deskę wylotową, która sięga do wewnątrz gołębnika i zaopatrzona jest na końcu w drewniany balast wyważany nogą gołębia stojącego na jej zewnętrznym końcu (rys. 88). Jeżeli na desce stanie szkodnik, który przeważnie jest cięższy niż gołąb, deseczka się złoży i zablokuje wejście. Gołębniki powinny być wykonane z dostatecznie grubych desek (18—24 mm), połączonych na pióro i wpust. Gołębnik musi mieć konstrukcję prostą i łatwą do rozbierania. Do jego budowy zamiast gwoździ powinno się używać wkrętów do drewna namoczonych w oleju lub nafcie, ewentualnie poszczególne ściany opatrzyć okuciami tak, aby gołębnik można było łatwo złożyć i rozebrać. Jest to konieczne do utrzymania w nim czystości i przeprowadzenia od czasu do czasu dezynfekcji. Ze względów higienicznych wewnętrzne strony drewnianych ścian powinny być ostrugane. Dobrze jest także, gdy ściany zewnętrzne są też ostrugane. Do budowy gołębnika wybiera się deski niezbyt wąskie, aby zmniejszyć liczbę szpar, w których się gnieździ robactwo. Nie mogą one być też zbyt szerokie, ponieważ wtedy bardziej się skręcają. Ozdabianie gołębnika nie jest wskazane. Utrudnia ono eksploatację gołębnika i zabiegi higieniczne, a efekt estetyczny przeważnie jest problematyczny. Dostęp do gołębnika musi być łatwy. W tym celu jedną ze ścian, najlepiej tę, do której jest najwygodniejszy dostęp, wykonuje się bądź jako drzwiczki otwierające się na bok, do góry lub na dół, bądź jako drzwiczki wyjmowane, umocowane haczykiem z oczkiem. Bardzo ważne jest urządzenie wnętrza gołębnika. Budkę dla każdej pary gołębi rozdziela się pośrodku deską 12 lub 15 mm grubości na dwie części (rys. 82). W jednej części są młode, a w drugiej gołębice na jajkach. Niektórzy hodowcy zalecają rozdzielać budkę — oczywiście dłuższą (ok. 1 m) — na trzy części. Obie boczne części są przeznaczone na gnieżdżenie się, a część środkową z otworem wylotowym tworzy przedsionek dla przylatujących i wylatujących gołębi. Gołębniki takie pokazano na rysunku 83 i 84. Przedsionek zaopatruje się w małe siodełko na przykład na tylnej ścianie, nad otworem wylotowym i na przegradzających deseczkach (jak listwa ułożona na płasko). Deseczki przegradzające budkę mają na całej długości wysokość 15 cm lub w przedniej części wysokość 15 cm, a w tylnej 20 cm. Deseczka przegradzająca spoczywa swobodnie na podłodze, oparta o przymocowane do niej dwie podpórki kątowe z grubej blachy albo konsolki z cienkich drewnianych listewek. Podpórki kątowe przymocowuje się do deseczki śrubą do drewna (wkręt do drewna) lub gwoździkiem. Podpórki kątowe mają otwory tylko na pionowych ramionach. Listewki osadza się w deseczce na wrąb dostosowany do profilu listwy. Aby gołębiom lepiej się siedziało na deseczce przegradzającej, przymocowuje się na jej krawędzi 4 cm listwę na płask.Rysunek 98 przedstawia gołębnik budkowy umieszczony na ścianie strychu, który jest o wiele lepiej dostępny niż gołębnik umieszczony na zewnętrznej stronie ściany. Otwór wylotowy może mieć wymiary od 12x15 do 15x16 cm, w zależności od wielkości gołębi. Wykonuje się go od 10 do 15 cm nad podłogą budki, aby nie dopuścić do wypadania młodych na zewnątrz. Przed wylotem, na zewnętrznej ścianie gołębnika, jest przymocowana deska wylotowa o grubości 1,8 cm, szerokości tej samej co szerokość otworu wylotowego albo trochę szersza i długości około 15 cm. Przeważnie deska ta jest podparta konsolką. Deska wylotowa może być też zamontowana wzdłuż całej długości budki, szerokość około 20 cm. Zamiast deski wylotowej przymocowuje się nieraz na spodnią deskę w otworze wylotowym listwę o szerokości około 2 cm, na górze nieco zaokrągloną albo osadza się na całej długości gołębnika deseczkę szerokości około 15 cm. Dla gołębi pocztowych desek wlotowych nie montuje się. Gołębniki dla cięższych gatunków gołębi nie powinny być umieszczone zbyt wysoko. Gołębie dobrze latające powinny mieć natomiast otwór wylotowy wyżej umieszczony. Jeśli obok siebie umieszcza się większą liczbę budek dla gołębi, oddziela się je na froncie pionowymi deseczkami o szerokości około 15 cm, aby gołębie stojące na deskach wylotowych nawzajem się nie widziały i nie przeszkadzały sobie (rys. 82). Dla każdej pary gołębi umieszcza się w budce jedno gniazdo (rys. 85, 86). Najlepsze są gniazda wystrugane z drewna oraz gniazda z mieszaniny gipsu i rozmoczonego papieru, ewentualnie z domieszką trocin (rzadką mieszaninę nalewa się do blaszanej formy wysmarowanej tłuszczem i po około 20 minutach po związaniu gipsu wyjmuje się gotowe gniazdo; pęcherze na powierzchni gniazda zasklepia się, a gniazdo wygładza zanurzając je w rozrobionym rzadkim gipsie). Nie należy wykonywać gniazd kamionkowych (są zimne) i słomianych (trzyma się w nich robactwo). Średnica gniazda wynosi od 19 do 34 cm, a wysokość od 4 do 8 cm i zależy od tego, dla jakiego gatunku gołębi jest ono przeznaczone. Bardzo ważna jest odpowiednia wielkość gniazda. Gniazda o podstawie czworokątnej są lepsze niż gniazda okrągłe, ponieważ po umieszczeniu ich w kącie nie zostaje za nimi wolna przestrzeń, gdzie mogą wpadać młode; są też stateczniejsze (rys. 86d). Gołębniki komorowe. Urządzamy je w budynkach na piętrze, na parterze lub w nadających się do tego pomieszczeniach na strychu, z których jest możliwy wylot w kierunku wschodnim, południowo-wschodnim lub południowym. Ściany i strop gołębników komorowych umieszczonych na strychu (rys. 87, 88, 89) wykonuje się najczęściej z desek (osłoniętych ewentualnie po zewnętrznej stronie gęstą drucianą siatką) albo izolacyjnych płyt budowlanych, np. wiórowo-cementowych, płyt drewna warstwowego, z obu stron gładko otynkowanych. Płyty i deski przybija się na szkielet z drewnianych krawędziaków, ewentualnie na słupy, kleszcze, krokwie i inne elementy więźby dachowej, jeśli mają odpowiednie położenie. Strop gołębnika komorowego musi być dobrze izolowany zarówno przed zimnem, jak i nagrzaniem w lecie. Z tego powodu stropem w gołębniku nie powinno być tylko pokrycie dachu, przez które w zimie łatwo przenika mróz, a które w lecie rozgrzewa się tak mocno, że w komorze gołębnika jest gorąco i duszno. Krokwie trzeba od spodu podbić deskami albo lepiej płytkami izolacyjnymi. Przestrzeń między krokwiami, pokryciem i podbiciem pozostawia się wolną. Jeśli strych jest dostatecznie wysoki, wykonuje się dla gołębnika strop (cały albo jego część, zależnie od okoliczności) samodzielny, poziomy, złożony z belek, pułapu, ewentualnie podsufitki. Zamiast desek można użyć płyt izolacyjnych zarówno na pułap, jak i na podsufitkę. Jeśli ściany gołębnika są murowane, nośne, wykonuje się strop z płyt ceramicznych lub prefabrykowanych żelbetowych płyt stropowych. Podłogę wykonuje się z cegieł, płytek ceramicznych z żużlobetonu z gładzią ceramiczną i z desek (z zakitowanymi szparami). Drewnianą podłogę dobrze jest pomalować farbą olejną albo jasnym karbolineum. Pod podłogę można włożyć, w celu zabezpieczenia przed myszami, gęstą siatkę drucianą. Wielkość gołębnika komorowego zależy od liczby gołębi — na jedną parę przewiduje się od 0,5 do 0,8 m2 powierzchni. Jeżeli projektuje się gołębnik mały, przyjmuje się normę większą i odwrotnie. Na przykład projektując gołębnik dla sześciu par gołębi należy przyjąć wymiary 6x0,8 —4,8 m2 powierzchni, tj. 2,00x2,40 m, w gołębniku dla 15—20 par gołębi hodowlanych należy przyjąć normę 0,5 m2 na 1 gołębia (komora dla 15 par gołębi powinna mieć wymiary 15x0,5 = 7,5 m2, tj. około 2,20x3,40 m). Aby można było oddzielić gołębie różnych płci, należy wyposażyć komorę w lekką, nieprzezroczystą, składaną przegródkę (na przykład z grubej papy albo lekkich płyt na szkielecie drewnianym). Umożliwia ona podzielenie komory na dwie części. O tym urządzeniu trzeba pamiętać już podczas budowy komory i odpowiednio rozmieścić gniazda, drzwi oraz otwór wylotowy. Wylot należy tak usytuować, aby również można go było rozdzielić przez środek przegródką na dwa otwory. W okresie zimowym jednym wypuszcza się jednego dnia samce, a następnego dnia drugim gołębice. Można także wykonać w gołębniku dwa wyloty, po jednym z każdej połowy komory. Każdej parze gołębi trzeba przydzielić jedno gniazdo o wymiarach: długość od 80 do 100 cm, szerokość i wysokość od 35 do 40 cm. Można umieszczać nad sobą aż cztery gniazda. Stawia się je przy bocznych i tylnych ściankach, tak aby środek komory pozostawał wolny, a promienie słoneczne nie padały bezpośrednio na gniazdo. Jeśli to możliwe, gniazd nie należy umieszczać naprzeciwko otworu wylotowego. Okna w gołębniku komorowym powinny stanowić od 1/6 do 1/8 ogólnej powierzchni podłogi. Powinny być one umieszczone nisko, około 30 cm nad podłogą, aby była ona dobrze oświetlona. Bardzo dobre są okna o podwójnych szybach. W lecie część okien zastępuje się ramą z gęstą siatką. Na wprost otworu wylotowego umieszcza się drzwi otwierane na zewnątrz. W drzwiach wykonuje się małe okienko obserwacyjne. Szerokość drzwi powinna wynosić 0,60 m. Drzwi powinny być szczelne, aby w komorze nie było przeciągu. Wysokość komory wynosi zwykle około 2 m. Wyższa komora nie jest dobra, ponieważ trudno wtedy chwytać gołębie; są one bardziej płochliwe. Na zewnętrznej ścianie, co najmniej jeden metr nad podłogą, znajduje się okienko wylotowe z wysuwanym lub odchylanym zamknięciem, o szerokości 60—80 cm i wysokości 35—40 cm. Okienko wyposażone jest również w opuszczaną albo wysuwaną drucianą bramkę. Aby nie przeszkadzać gołębiom znajdującym się w komorze, okienko otwiera się linką na blokach lub krążkach sterowanych ze strychu ewentualnie z zewnątrz, zwłaszcza jeśli okienko wylotowe znajduje się w szczycie budynku pod okapem. Szczelnych zamknięć używa się w zimie, drucianych bramek w lecie. Na zewnętrznej i wewnętrznej stronie otworu wylotowego umocowuje się, np. na konsolkach, deseczki szerokości około 30 cm. Wewnętrzna deseczka, na którą gołębie stają podczas opuszczania komory, jest nieco nachylona do wewnątrz, podczas gdy zewnętrzna deska wylotowa (lotka), na której gołębie po przylocie przysiadają, jak również chętnie przechadzają się i rozglądają, jest nachylona (około 1 cm) na zewnątrz. Ze względu na konieczność przegrodzenia komory w pewnych okresach hodowlanych zarówno rama okienka wylotowego, jak i drewniana bramka są dwuskrzydłowe, aby obie połowy mogły się samodzielnie otwierać i zamykać. Pożądane jest, aby w bezpośrednim sąsiedztwie lub niedaleko gołębnika znajdował się np. gzyms lub dach o nieznacznym nachyleniu na niższym budynku, aby gołębie mogły z nich zlatywać do gołębnika. Na bardziej stromym dachu można w tym celu zamocować jedno lub więcej siodełek z desek, osadzonych na metalowych podpórkach w dachu. W otworze wylotowym można także umieścić tzw. zamknięcie automatyczne (rys. 90). Najczęściej są to dwa poziome druty przeciągnięte w poprzek otworu wylotu. Na górnym drucie zawiesza się szereg prętów, które wolno opadają ku dołowi aż do oparcia się o niższy drut poziomy. Pręty można odchylić tylko w jedną stronę, np. do wnętrza pomieszczenia, co uniemożliwia ptakom swobodne wychodzenie z gołębnika. Pręty mogą być też zainstalowane w taki sposób, aby połową otworu wylotowego gołębie mogły wychodzić z gołębnika, a połową wchodzić. Wielu hodowców umieszcza u wylotu tzw. skrzynkę startową (lotkę rys. 91, 92). Ściany i sufit skrzynki startowej wykonane są z drutu lub siatki drucianej rozciągniętej na szkielecie z listew lub prętów stalowych. Środek ściany szczytowej lub ściany bocznej stanowią drzwiczki otwierane przeważnie za pomocą linki (sznura) ze strychu albo z zewnątrz. Podłoga skrzynki wykonana jest z grubych desek wystających poza ściany boczne na długość 50—60 cm lub tworzących rodzaj ławeczki biegnącej wzdłuż ściany frontowej. Jeśli budynek nie ma podmurówki strychowej (ścianki tremplowej) albo jest ona za niska, otwór wylotowy gołębnika umieszcza się bezpośrednio na ścianie obwodowej, nadbudowując ją do odpowiedniej wysokości (rys. 87). Szczytowa ścianka z otworem wylotowym może być także wymurowana. W komorze gołębnika umieszczonej z dala od ścian strychowych otwór wylotowy wykonuje się w dachu. Taki otwór wylotowy musi być także zabezpieczony przed szkodnikami. Na dachu nad wylotem można wykonać także skrzynkę startową z ocynkowanej siatki drucianej (rys. 93), tzw. wolierę. Komorę należy dobrze wietrzyć, oczywiście nie wywołując przeciągu. W mniejszych komorach wietrzenie odbywa się głównie poprzez otwór wylotowy. W większych gołębnikach do wietrzenia służą oprócz wylotu jeszcze otwory okienne. Okna są na ogół także potrzebne do dostatecznego oświetlenia komory. Zaopatruje się je w drucianą siatkę. Niekiedy wykonuje się jeszcze małe otwory w drzwiach i w stropie, ewentualnie także w zewnętrznej ścianie, zakryte gęstą drucianą siatką. Ściany i strop komory należy pobielić. Gniazda w gołębniku komorowym pokazano na rysunkach 94, 95, 96, 97, 98, 99 i 100. Należy zawsze pamiętać o tym, aby gniazda można było łatwo oczyścić i rozbierać. Bardzo dobre są gniazda umieszczone w drewnianych albo stalowych stojakach lub z wyjmowaną przynajmniej jedną — przednią — ścianą oraz przegrodą między obydwoma przedziałami. Długość gniazda wynosi od 85 do 100 cm, głębokość 40 cm i wysokość od 35 do 40 cm. Zwykle umieszcza się po trzy gniazda nad sobą. Każde gniazdo przegradza się w środku wyjmowaną przegródką na dwie części, jedną — przeznaczoną do wysiadywania jaj, drugą — dla dorastających młódków. Przegródka może ewentualnie sięgać aż do sufitu gniazda. Część przedniej ściany gniazda z reguły wykonuje się z listew albo z drutu średnicy około 3 mm. Jeśli przy gniazdach umocuje się deskę wylotową, można użyć jej jednocześnie do zamknięcia gniazda, a to przez podniesienie jej w górę, do otworu wylotowego (rys. 94). W innym wypadku gniazdo zamyka się według potrzeby lekką bramką, zawieszoną lub przesuwającą się na poziomych prowadnicach. Deska powinna być tak wykonana, aby można ją było wyjąć w celu oczyszczenia. Z tego samego powodu również podłoga gniazd powinna być wysuwana. Jeśli deski wylotowe umieszczamy wzdłuż całej długości gniazda (przy wszystkich gniazdach lub tylko przy co drugim), trzeba oddzielić pionowe rzędy gniazd przegrodą. Na ścianie gołębnika komorowego trzeba koniecznie umocować siodełka dla gołębi, zwłaszcza młódków. Ulubione przez gołębie są pojedyncze lub umieszczone na pionowej listwie jedno nad drugim siodełka przyścienne daszkowe wykonane z deseczki o szerokości około 7 cm i długości 15 cm. Siodełka mają po obu stronach ukośne „skrzydełka” z blachy ewentualnie deseczek, długości od 25 do 30 cm, które chronią niżej siedzące gołębie przed zanieczyszczeniem odchodami (rys. 101). Oba skrzydełka wykonane są z jednego kawałka blachy, który przechodzi przez wycięcie w deseczce służącej do siedzenia. Aby ułatwić zatrzymywanie odchodów na deseczkach tworzących skrzydełka, oba końce skrzydełek są zagięte do góry. Siedzenia tarczowe - z siodełkami talerzowymi o średnicy około 15 cm są przeznaczone głównie dla garłaczy i pawików. Siedzenia zespołowe z deseczki o szerokości od 8 do 10 cm, są przegrodzone co około 20 cm drewnianymi deseczkami, aby gołębie nawzajem sobie nie przeszkadzały. Pod siodełkami umieszcza się deseczkę służącą do zatrzymywania odchodów. Siedzenie zespołowe przedstawiono na rysunkach 102 i 103. Do wyposażenia gołębnika komorowego należy jeszcze korytko na składniki mineralne paszy, poidło na czystą wodę i naczynie do kąpieli. Korytko powinno być tak wykonane, aby gołębie nie mogły do niego wchodzić i zanieczyszczać go. Dlatego od góry zamyka się je klapą, a w otworach, umieszczonych w bocznych ścianach zakłada się kratkę z ukośnych albo pionowych grubych drutów lub zaokrąglonych listew (np. z prętów leszczynowych) w odstępach 5—6 cm (większych dla gołębi rasy dużej). Podczas karmienia każdy gołąb powinien dziobać tylko w tym otworze, który zajął. Obustronne karmidła pokazano na rysunku 104 i 105. Karmidło na rysunku 104 ma w środku wzdłuż długości ukośne deseczki, które nie pozwalają dwom gołębiom dziobiącym na przeciw siebie przeszkadzać sobie nawzajem. Ukośne deseczki umocowane co 1—1,5 cm pozwalają na równomierne rozłożenie ziarna na dnie korytka. W korytkach dla gołębi z pierzastymi ozdobami na głowie i szyi oraz dla garłaczy nie daje się krat. Korytko odpowiednie przede wszystkim dla garłaczy pokazano na rysunku 105. Jest ono wyposażone w pionowe przegródki, które uniemożliwiają gołębiom widzenie się nawzajem podczas pobierania karmy. Dzięki temu gołębie podczas karmienia nie odzierają sobie woli. Samoczynne karmidło pokazano na rysunku 106. Karmidło na mineralne domieszki pokazano na rysunkach 107 i 108. Są to skrzyneczki z dnem przechylającym się w kierunku przedniej ściany, z otworami do karmienia o szerokości 5—8 cm. Otwory zamyka się blaszaną zastawką. Karmidło od góry jest zakryte klapą. Do pojenia gołębi, które potrzebują stosunkowo dużej ilości wody, używa się prostych poideł kamionkowych, wykonanych z żeliwa lub emaliowanych, o wysokości od 5 do 6 cm. Można stosować także poidła samoczynne, z blachy, butelki itp, które pokazano na rysunkach 109 i 110. Poidła powinny być tak wykonane, aby się w nich gołębie nie mogły kąpać i nie zanieczyszczały ich łajnem. Karmidła i poidła najlepiej jest umieścić na ławeczce lub niskim stoliku. Naczynie do kąpania wykonuje się z blachy emaliowanej albo ocynkowanej, ewentualnie z kamionki, o wielkości 40x50 cm i wysokości 5—6 cm, z szerokim brzegiem (rys. 111). Napełnia się je wodą na wysokość 4—7 cm. Do wyposażenia gołębnika komorowego należy jeszcze klatka godowa przeznaczona dla jednej pary gołębi. Jest ona tak duża jak gniazdo i ma ściany i sklepienia wykonane z drutu. Klatka rozdzielona jest pionową zasuwaną przegródką z drutów osadzonych pionowo w ramce (rys. 112). W obu połowach przedniej ściany, którą można wyjmować, ponieważ jest umocowana na zakrętkach, znajdują się małe druciane drzwiczki. Klatka godowa służy także do tresury gołębi przeznaczonych na wystawę. Gołębniki domowe. Jeśli w pobliżu domu jest dostateczna ilość miejsca, bardziej wymagający hodowcy stawiają samodzielne domki dla gołębi (rys. 113), połączone często z ogrodzonym miejscem służącym do wylotów. Takie gołębniki są szczególnie odpowiednie do hodowli gołębi rzeźnych Nie zaleca się budować domków większych niż dla 20 par gołębi (bójki, większa możliwość infekcji itp.). Ściany domków mogą być wykonane z drewna, cegieł — najlepiej dziurawki, kształtek lub płyt konstrukcyjnych czy izolacyjnych, osadzonych na drewnianym szkielecie. Fundamenty domków drewnianych wykonuje się z betonu 1 : 8 o szerokości od 20 do 30 cm lub z cegieł o szerokości 30 (25) cm. Wystarczą także tylko filary fundamentowe o przekroju 30x30 cm. Drewniane ściany wykonane są ze szkieletu z krawędziaków i obite deskami (w miejscach osłoniętych z jednej strony, w miejscach wystawionych na działanie wiatru i położonych w wyższych rejonach z dwóch stron). Dach należy pokryć papą lub eternitem. Do wietrzenia, które jest bardzo ważne, służą okna i zabezpieczone siatką otwory w ścianach. Na zimę zakłada się do nich oszklone ramy. Okna umieszcza się w południowo-wschodniej, południowej albo wschodniej ścianie domku. Budując domek trzeba pamiętać o tym, aby można go było przesunąć lub przenieść w inne miejsce. Wewnątrz domek jest urządzony tak, jak gołębnik komorowy na strychu. Gniazda umieszczone są przy jednej lub dwu ścianach domku z wylotami prowadzącymi do wewnątrz pomieszczenia lub bezpośrednio do skrzynki startowej, jeśli ta z nimi sąsiaduje. W domku robi się tylko jeden wylot do skrzynki startowej (woliery), czasem podwójny, tzn. z dwoma otworami umieszczonymi obok siebie, aby wychodzące i wchodzące gołębie nie spotykały się. Drzwi do gołębnika powinny zamykać się samoczynnie. Skrzynka startowa (woliera) wykonana jest z cynkowanej siatki drucianej rozciągniętej na szkielecie z drewnianych, betonowych albo stalowych słupków (rurek). W górnej części klatki zastępuje się zazwyczaj siatkę daszkiem, aby gołębie mogły przebywać w skrzynce startowej również w czasie niepogody Skrzynka startowa przeznaczona dla 10 par gołębi ma powierzchnię około 6x4 m. Wysokość jej wynosi od 2,20 do 3,00 m. Siatkę należy wpuścić do ziemi na głębokość około pół metra. W ten sposób nie dopuści się do podkopywania się pod nią szkodników. Podłoga skrzynki startowej powinna być umieszczona tak, aby nie zatapiała jej woda. Powinna być ona wysypana czystym piaskiem. Zaleca się pod piasek dać podłoże betonowe o grubości około 8 cm. W razie wystąpienia choroby zaraźliwej piasek należy wyrzucić, a podłoże betonowe umyć roztworem dezynfekcyjnym i na nowo pokryć czystym piaskiem. Nie należy pozostawiać dna trawiastego. Ściany klatki osłania się u dołu za pomocą desek lub blachy na wysokość 30—40 cm od podłogi. Ze skrzynki startowej można także zrobić wylot na zewnątrz. Drzwi do skrzynki startowej prowadzą albo z gołębnika, albo z dworu. Nie należy ich umieszczać na przeciwko wylotu z domku. Wewnątrz skrzynki umocowuje się na ścianach deseczki do siedzenia lub pojedyncze siodełka. Aby gołębie nie mogły siadać na poziomej belce szkieletu, górną powierzchnię belek umieszczamy w położeniu ukośnym. Wokół ogrodzenia można zasadzić krzewy, pnącza i kwiaty. Autor: Vaclav Tajowsky Dla potrzeb portalu wedlinydomowe.pl opracował Maxell






















-
Rozumiem, że masz na myśli taki "nożyk"?

-
[Porady dla poczatkujących] Dyskusja
Maxell odpowiedział(a) na Muski temat w Podpowiadamy jak zrobić pierwsze wyroby
Podbijam temat nadsyłania na moja pocztę dokładnych fotorelacji (najlepiej krok po kroku) z odpowiednim opisem, na wymienione wyżej przetwory. Macie tego sporo, ale jak pisałem, nie mogę pobierać z forum. Poza tym, jest okazja by je uzupełnić zgodnie z obecną wiedzą. -
[Porady dla poczatkujących] Dyskusja
Maxell odpowiedział(a) na Muski temat w Podpowiadamy jak zrobić pierwsze wyroby
Nie poda zamienników, gdyż na wątrobiankę z mięsa wieprzowego będzie następny przepis. -
Izolacja wędzarni - wątek ogólny
Maxell odpowiedział(a) na Tedi19791 temat w Wędzarnie. Budowa i obsługa
Czy tylko w wędzeniu? -
Prezentuję Wam wątrobiankę produkcji Kolegi Arkadiusza: Wątrobianka grodziska z dzika i sarny z dodatkiem wieprzowiny A. Surowiec mięsno - tłuszczowy: 1. Mięso z kości (dzik, sarna) - 3 kg 2. Wątroba wieprzowa + wątroby z dzika i sarny - 3 kg 3. Podgardle wieprzowe - 3 kg 4. Skórki wieprzowe 70 dkg 5. Opcjonalnie rosół z gotowania mięs zależnie od tego czy ma być bardziej lub mniej smarowna Surowiec nie solony i nie peklowany. B. Przyprawy i materiały pomocnicze: I. Przyprawy 1. Sól – 17 g/kg 2. Pieprz - 0,5 g/kg 3. Majeranek - 1,5 g/kg 4. Cebula suszona – 3 g/kg Cebulę suszoną można zastąpić cebulą świeżą w stosunku 0,15 kg suszu - 1 kg cebuli świeżej. Cebulę świeżą kroimy w piórka i obsmażamy. II. Materiały pomocnicze 1. Jelita grube (odcinki proste) 2. Przędza C. Postać surowca po obróbce Wszystkie składniki rozdrobnione na wilku (maszynka do mięsa) przez siatkę nr 2 (średnica oczka to 2 mm) i kutrowane (rozdrabniane na kutrze lub blenderem). Wszystkie składniki (z wyjątkiem wątroby) przed rozdrobnieniem gotowane. D. Kiszka wątrobiana grodziska - skrót instrukcji 1. Solenie: Surowiec do produkcji powinien być nie peklowany i niesolony. Przy zastosowaniu surowca solonego należy zmniejszyć ilość soli dodanej w czasie produkcji. 2. Mycie i moczenie: Surowiec przed gotowaniem myje się 3. Gotowanie: Surowiec (z wyjątkiem wątroby) gotuje się w temp.95oC do miękkości. Podgardle w temp.85 C do stanu półmiękkiego. Po ugotowaniu kości obiera się z mięsa. 4. Rozdrabnianie: Zgodnie z recepturą 5. Kutrowanie: Rozdrobniona wątrobę kutruje się do stanu uzyskania mazistej masy, następnie dodaje się przyprawy, resztę ugotowanego zmielonego mięsa i podgardle i kutruje do wymieszania wszystkich składników. Ja mielę mięso trzykrotnie na sitku nr 2, dodaję wykutrowaną wątrobę, przyprawy i mieszam. Smarowność reguluję ilością dodanego rosołu z gotowania mięs. 6. Napełnianie: Wykutrowaną masą napełnia się dość luźno jelita. Końce wiążę się przędzą robiąc z jednej strony pętelkę do zawieszenia. Długość batonów nie powinna być zbyt duża, żeby w trakcie podwędzania się nie urwały. Ja robię batony długości ok. 15-20 cm. 7. Gotowanie: Kiszki wrzuca się do wrzącej wody i gotuje w temp.80-85oC do uzyskania wewnątrz kiszki temp. 68-70oC. 8. Studzenie: Kiszki po parzeniu studzi się przez 10 min w zimnej wodzie a następnie rozwiesza i studzi do temp. 12oC. 9. Podwędzanie: Po wystygnięciu kiszki podwędza się w temp. 35-40oC do uzyskania odpowiedniego koloru. Smacznego: Autor: Arkadiusz











-
[Porady dla poczatkujących] Dyskusja
Maxell odpowiedział(a) na Muski temat w Podpowiadamy jak zrobić pierwsze wyroby
Ale są w kąciku. Abym je wkleił, Arek musiałby mi podesłać oryginały zdjęć i tekst.