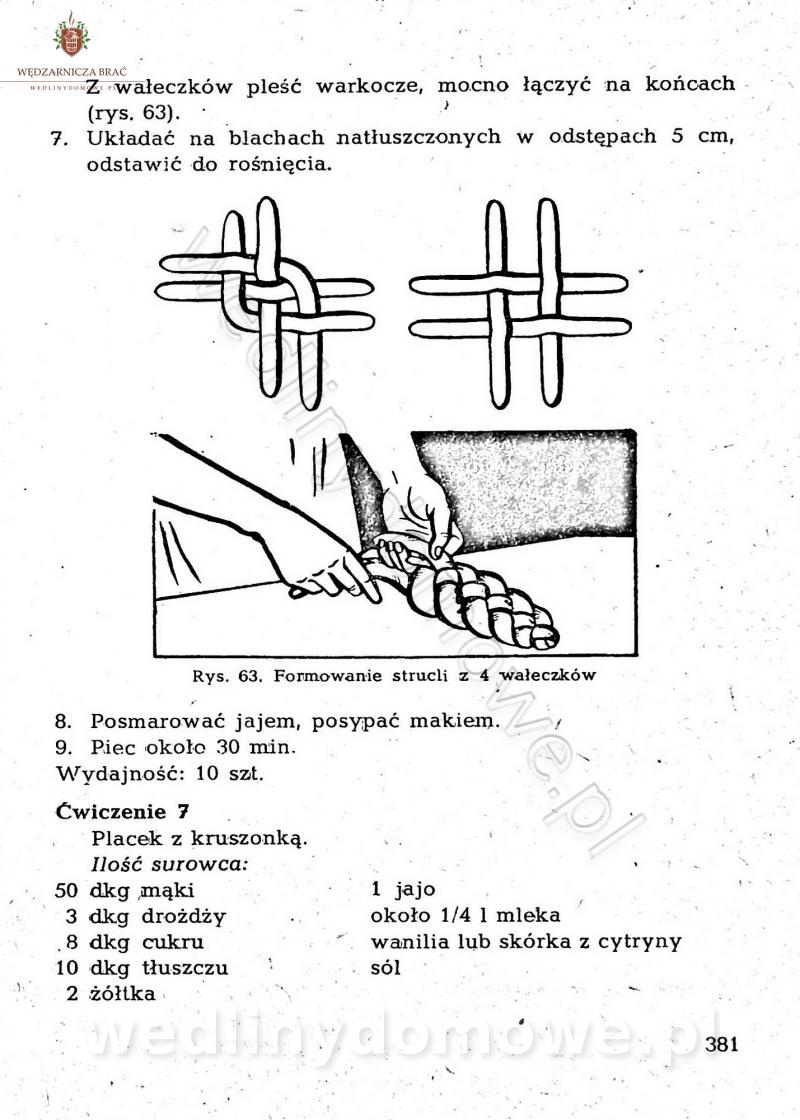
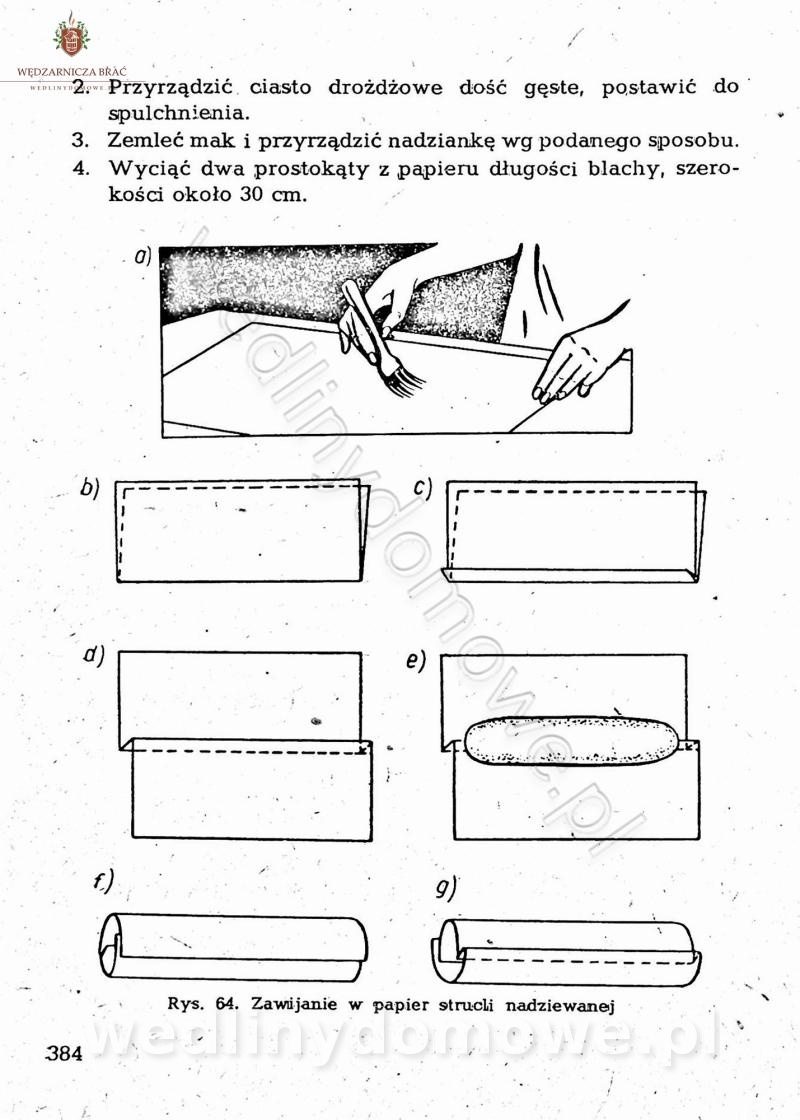
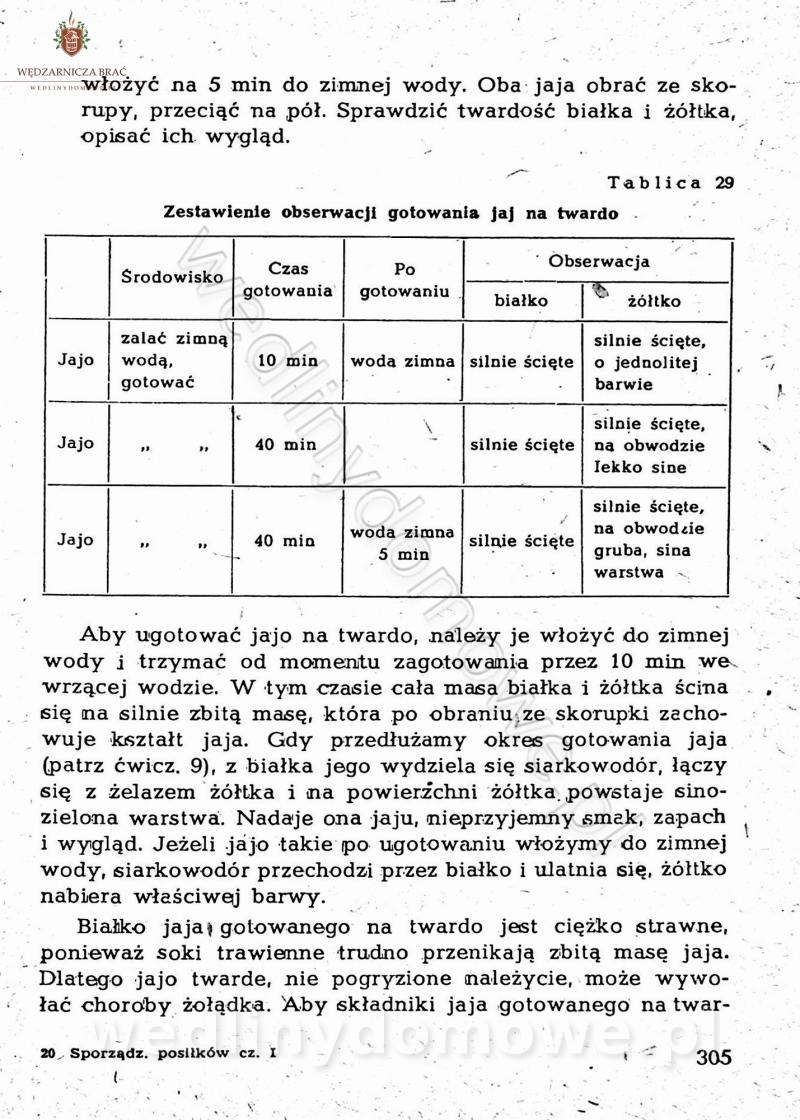
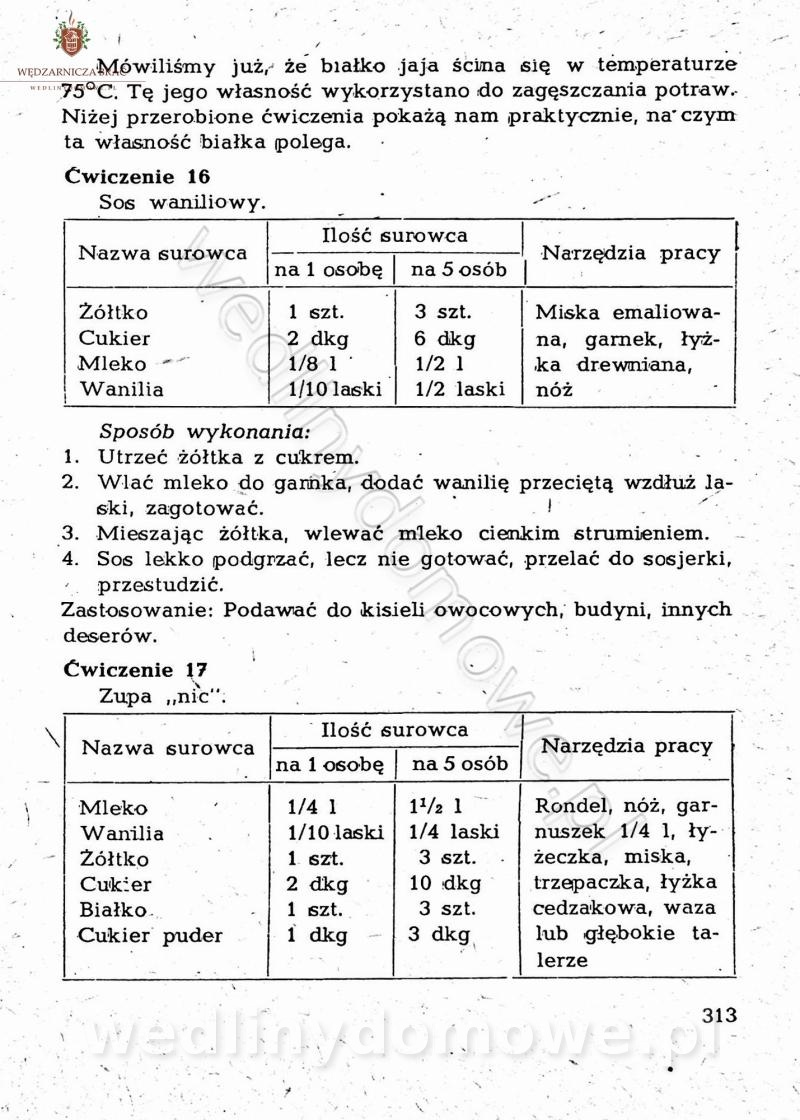
Maxell
-
Postów
46 204 -
Dołączył
-
Ostatnia wizyta
-
Wygrane w rankingu
180
Typ zawartości
Profile
Forum
Wydarzenia
Mapa użytkowników
Sklep
Giełda
Treść opublikowana przez Maxell
-
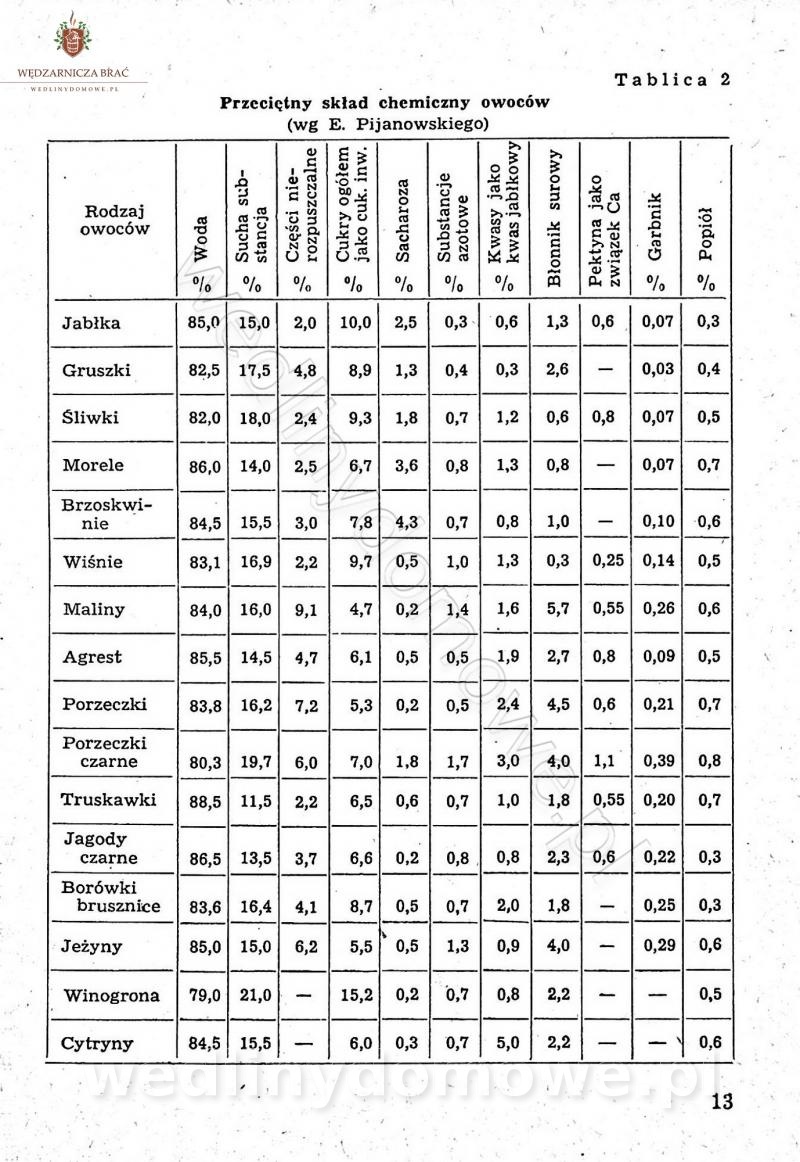
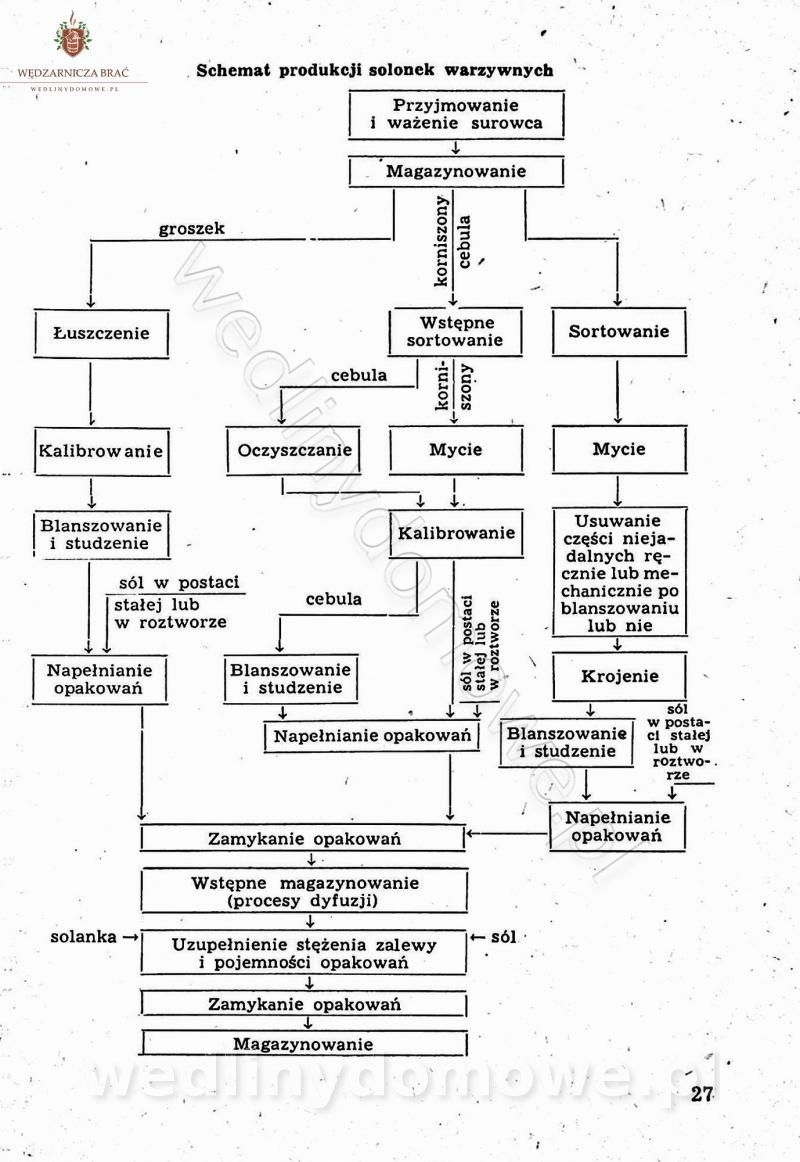
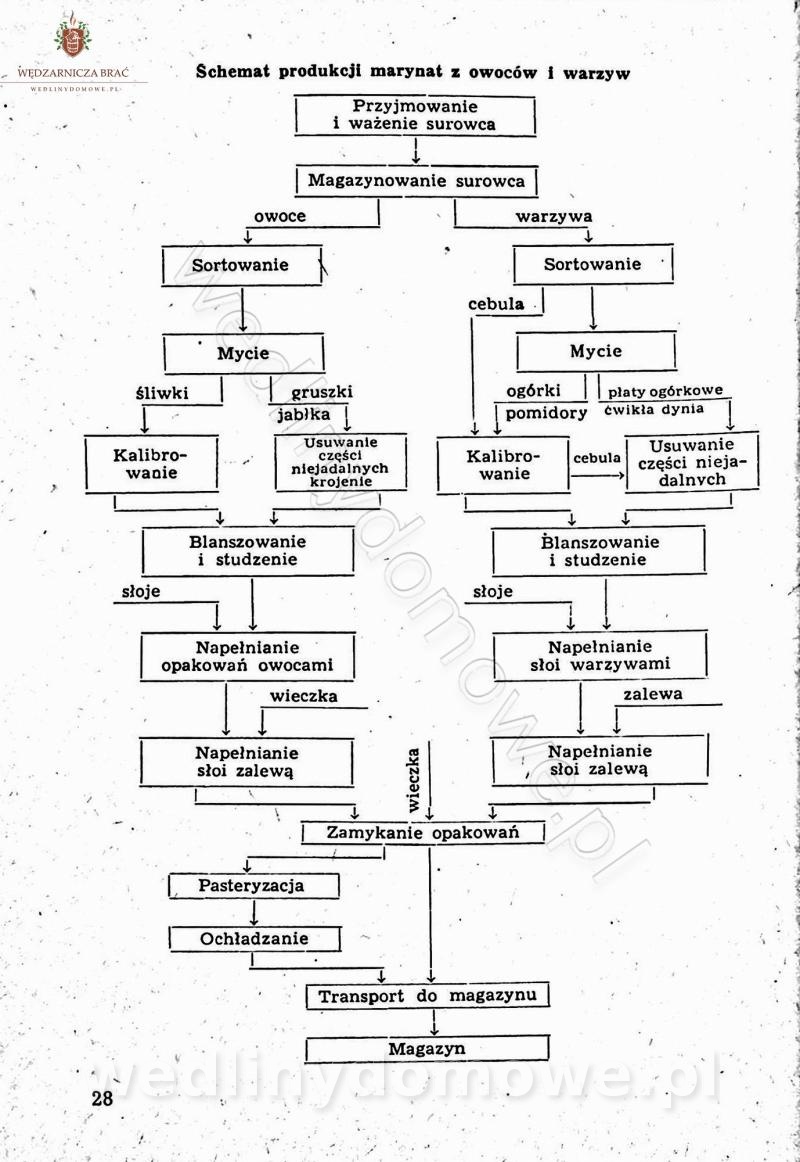
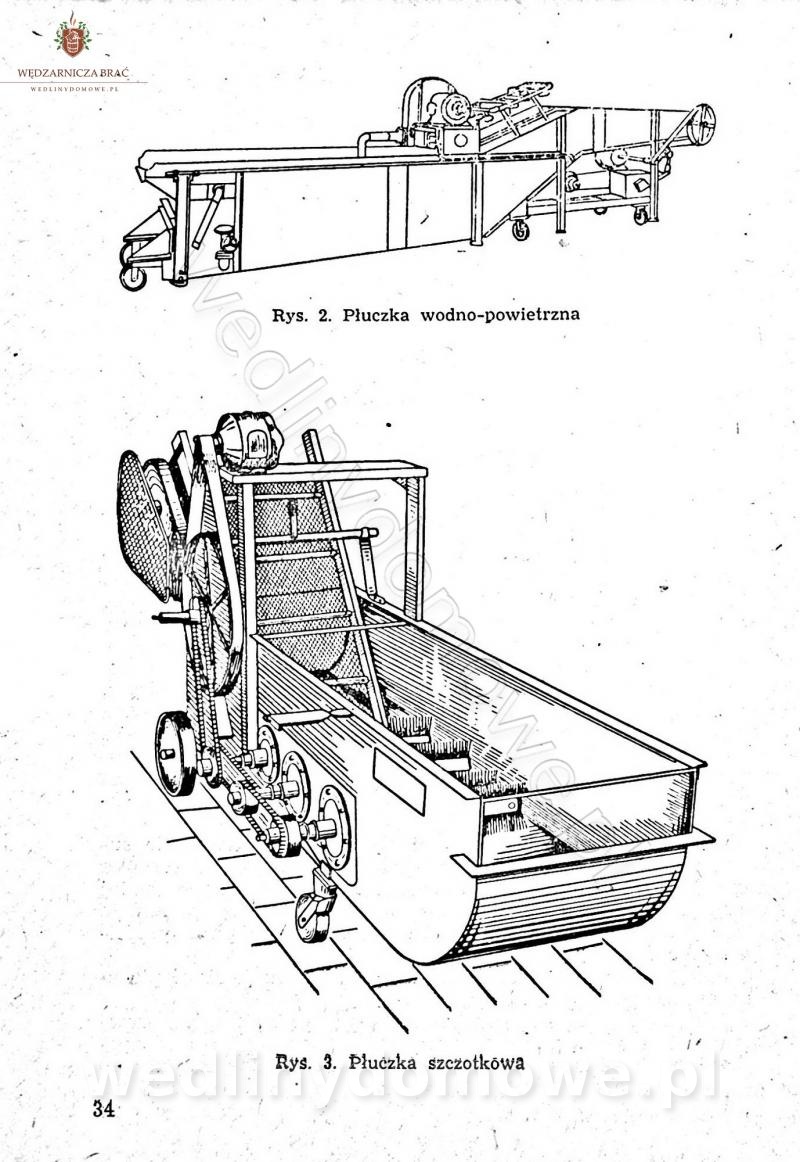
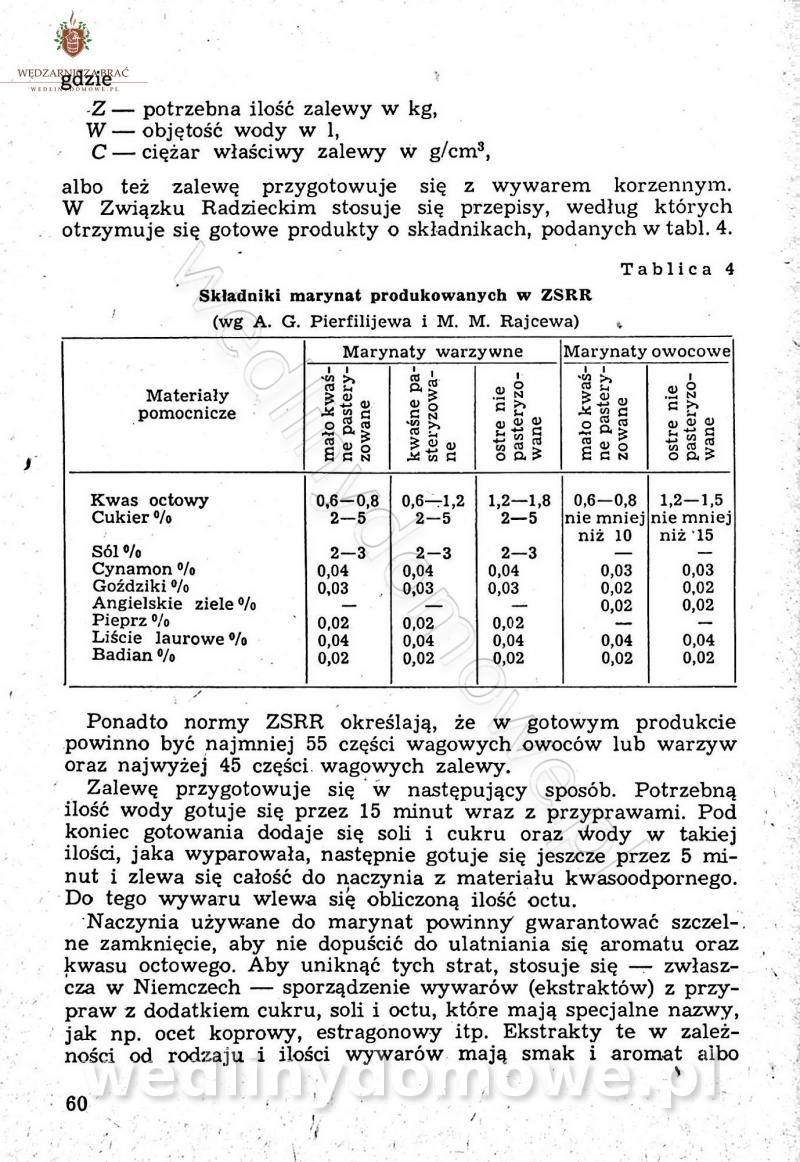
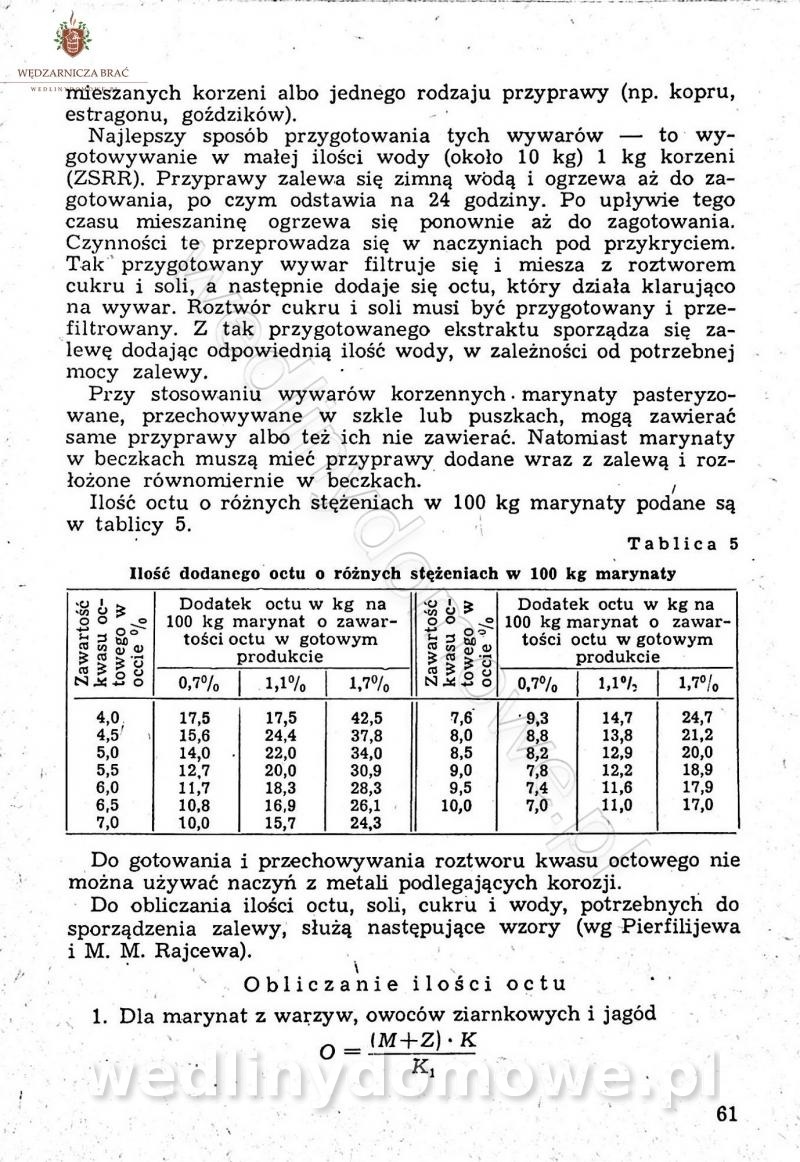
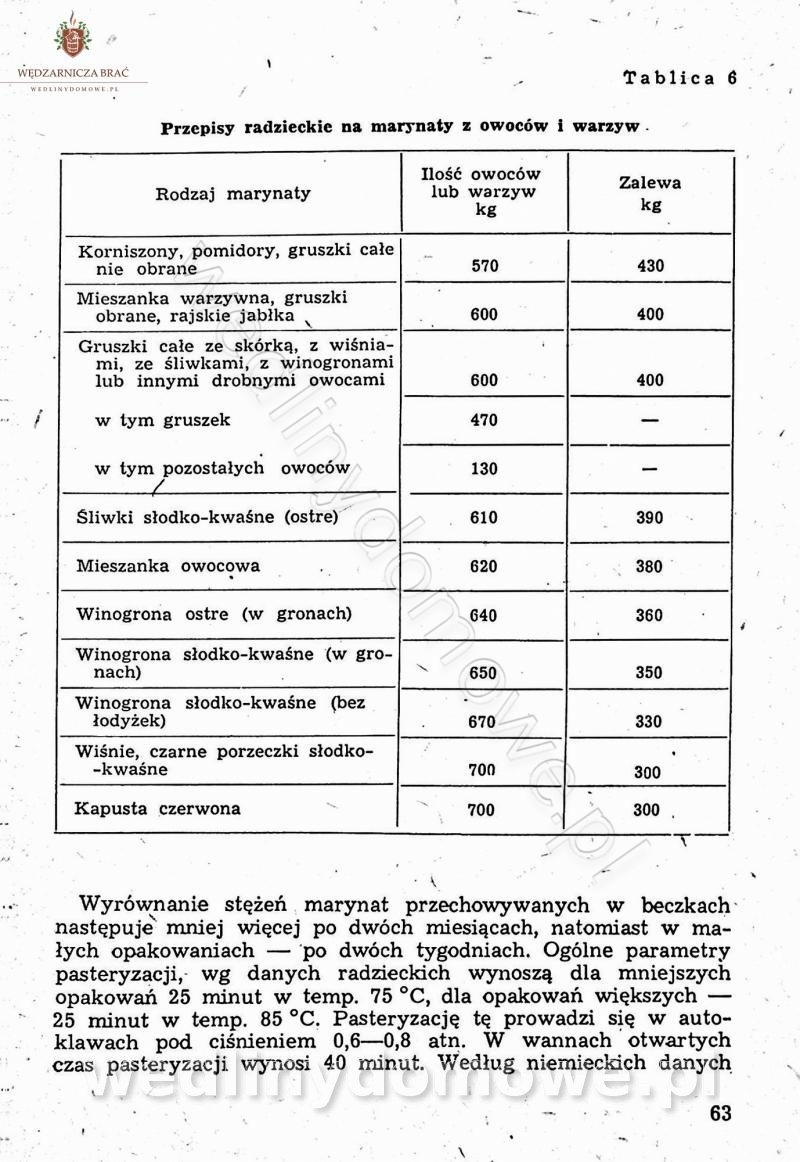
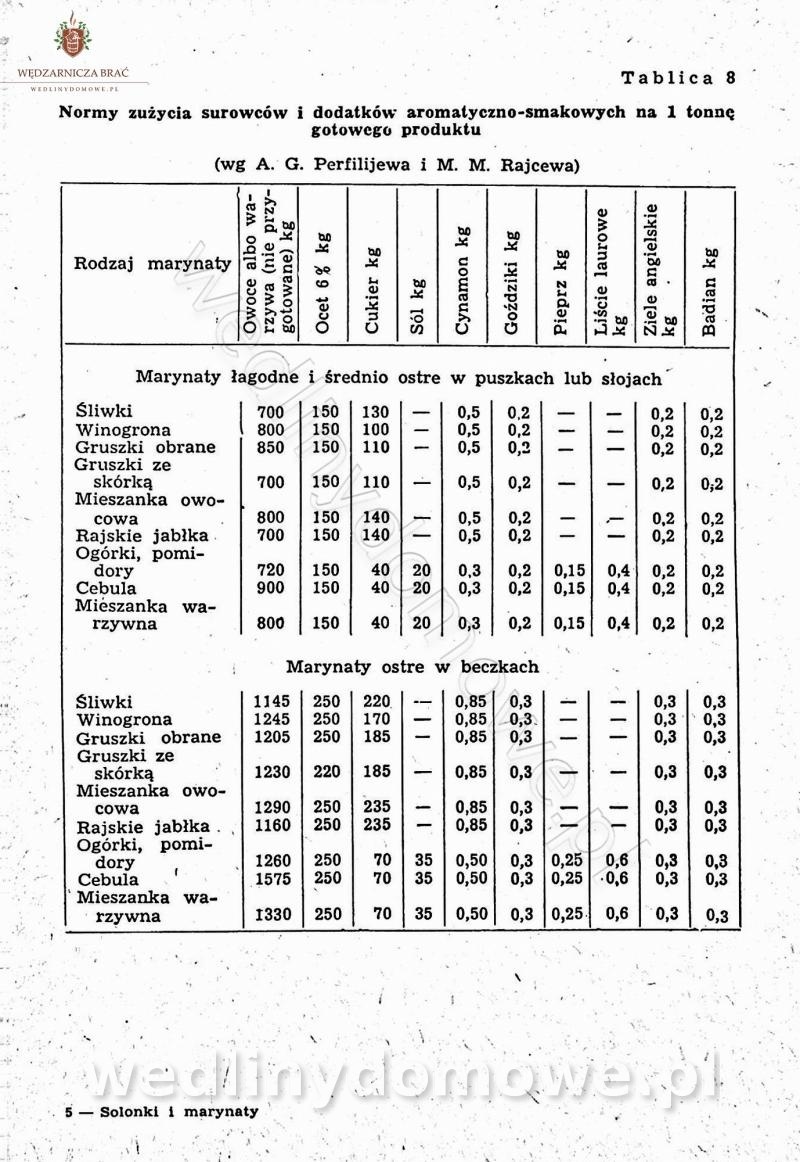
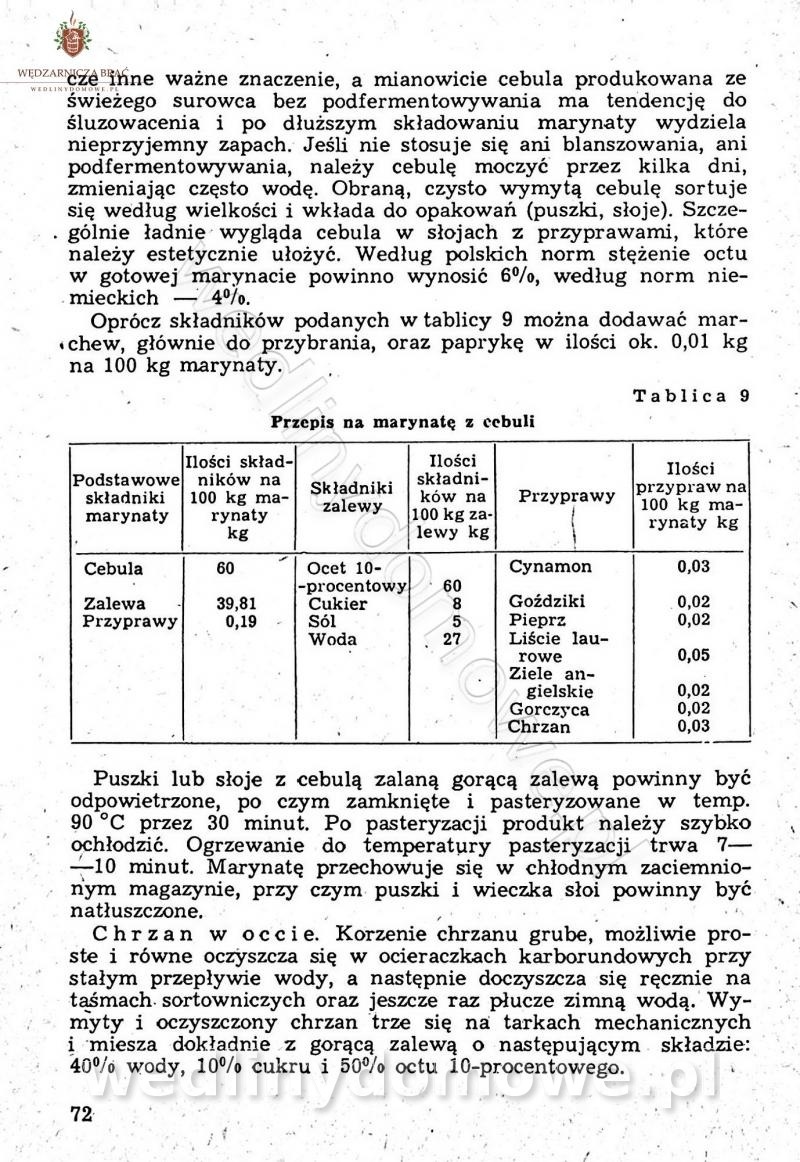
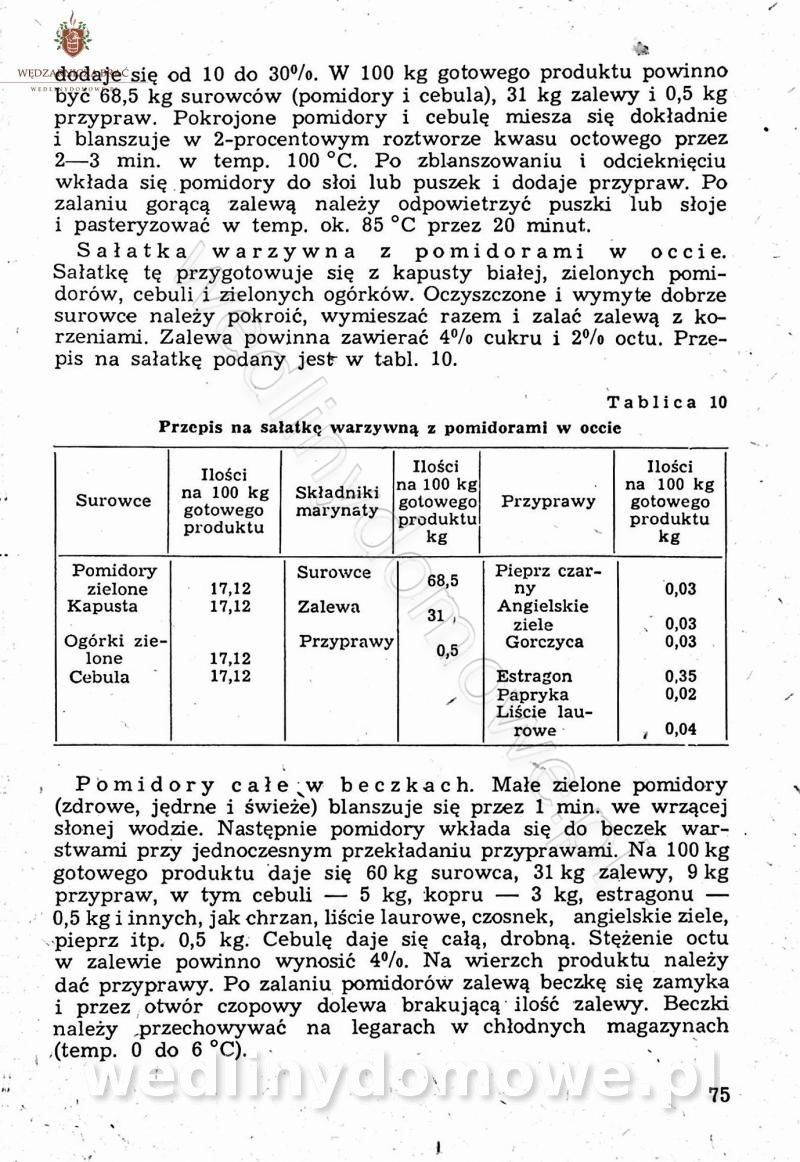
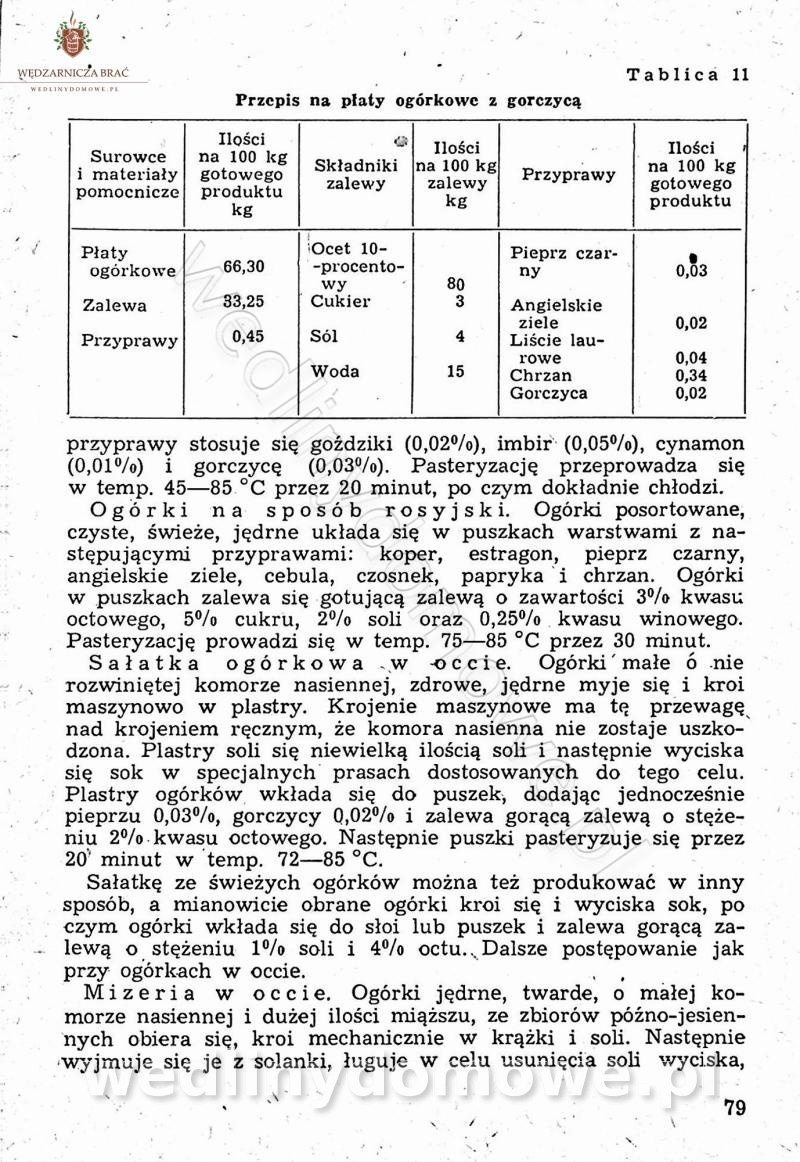
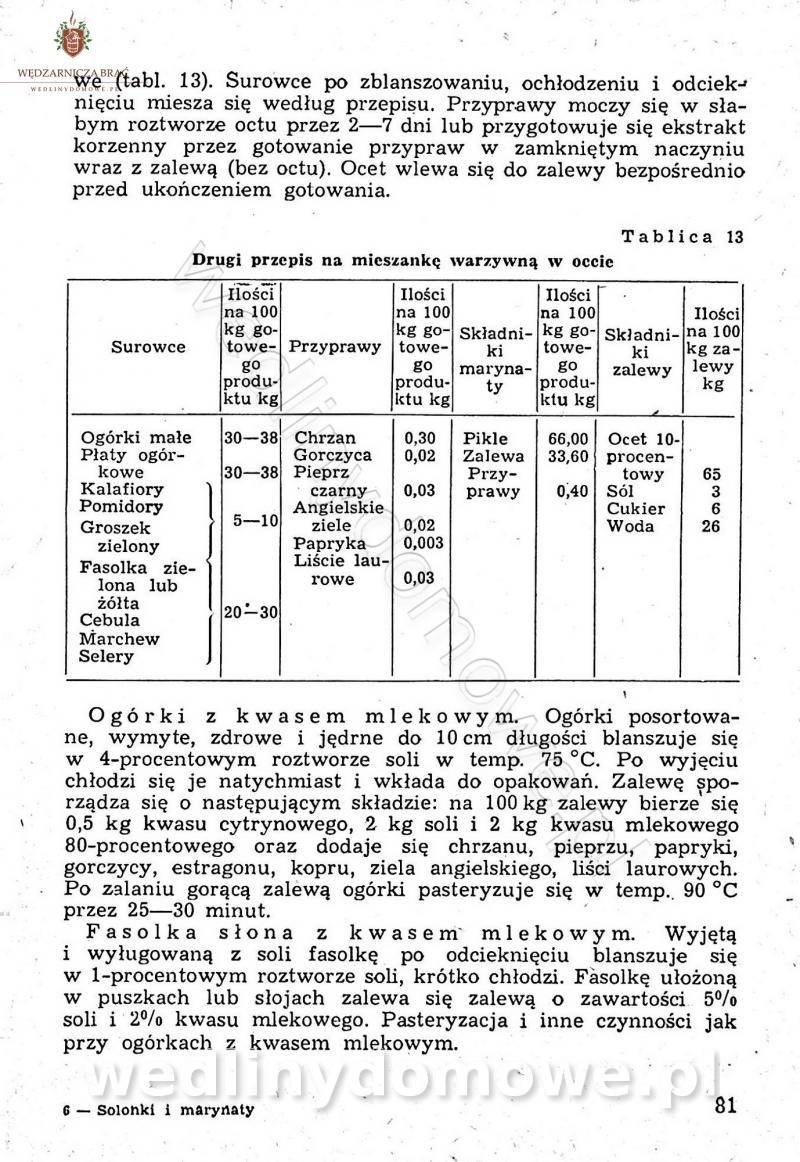
Polecam [Rok 1957] Solonki i marynaty z owoców i warzyw.
Maxell odpowiedział(a) na Maxell temat w Przetwórstwo owoców i warzyw
W przepisach występuje przyprawa o nazwie badianek. To nic innego, jak anyż gwiaździsty. -
Polecam [Rok 1957] Solonki i marynaty z owoców i warzyw.
Maxell odpowiedział(a) na Maxell temat w Przetwórstwo owoców i warzyw
Na potrzeby portalu wedlinydomowe.pl opracował Maxell
































-
Polecam [Rok 1957] Solonki i marynaty z owoców i warzyw.
Maxell opublikował(a) temat w Przetwórstwo owoców i warzyw
Długo szukałem tej książki, gdyż uważam że jest bardzo cenna.












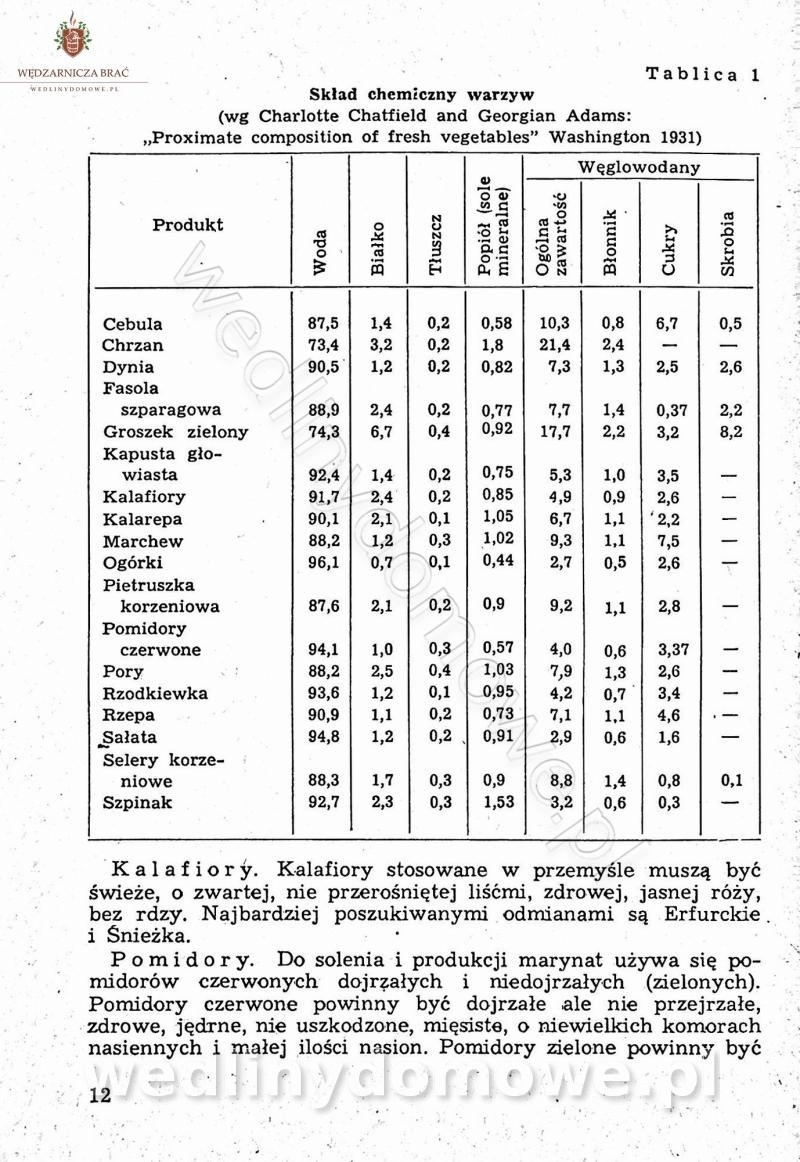
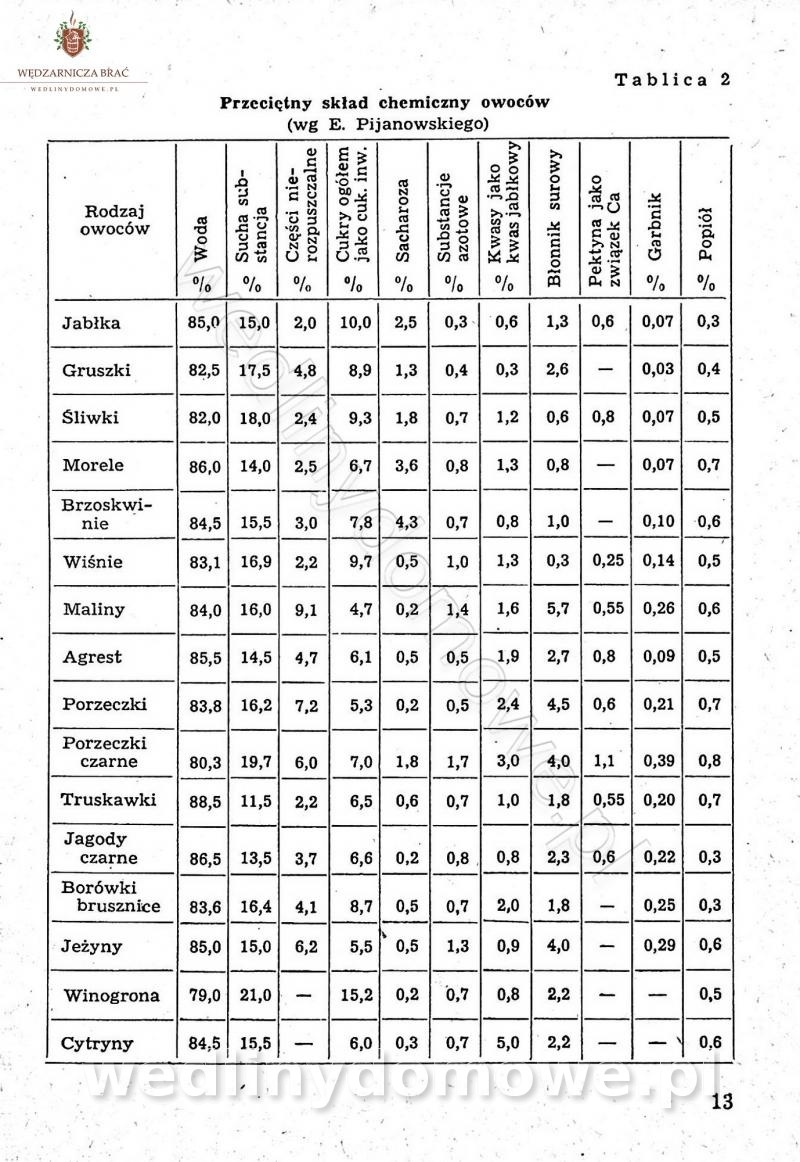













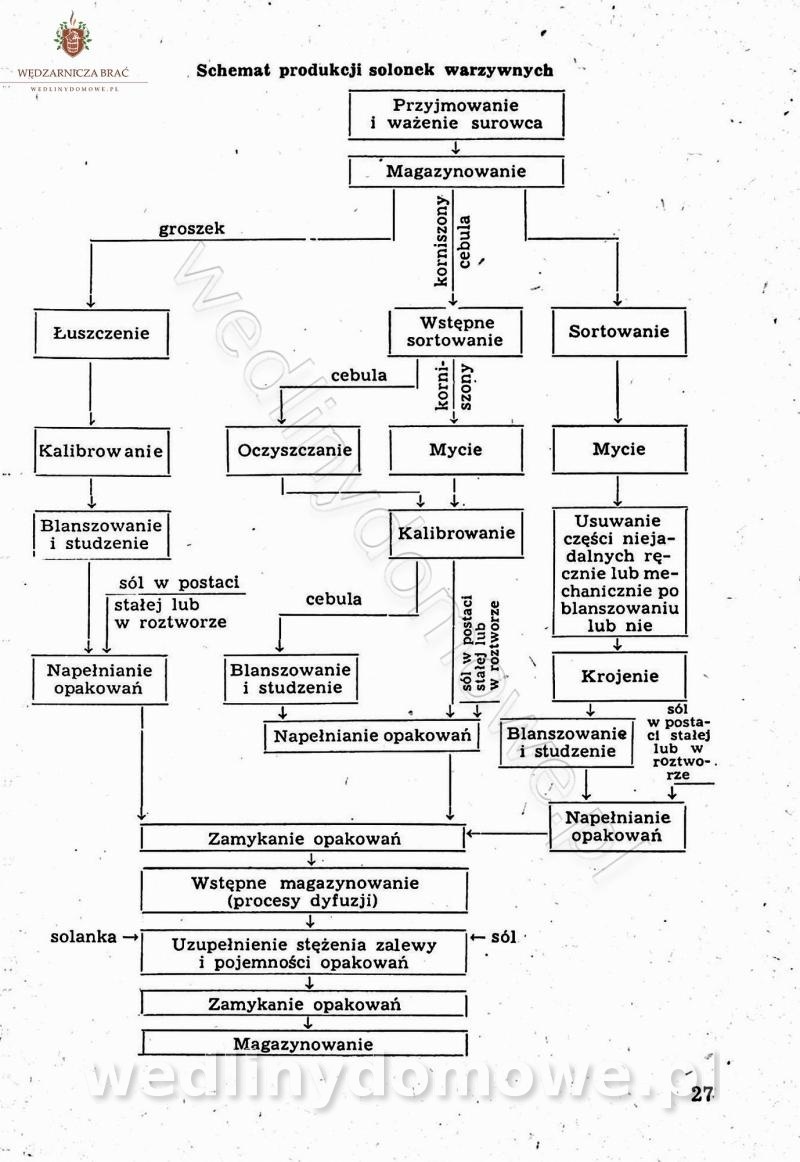
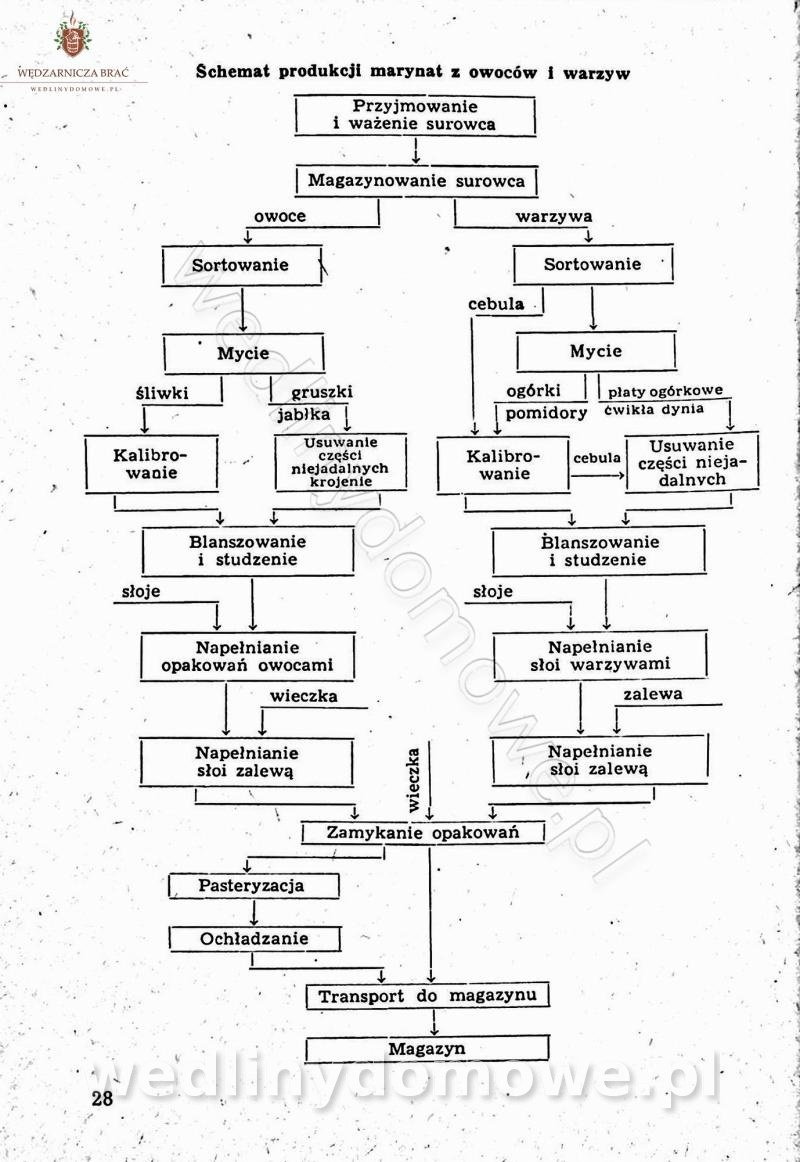





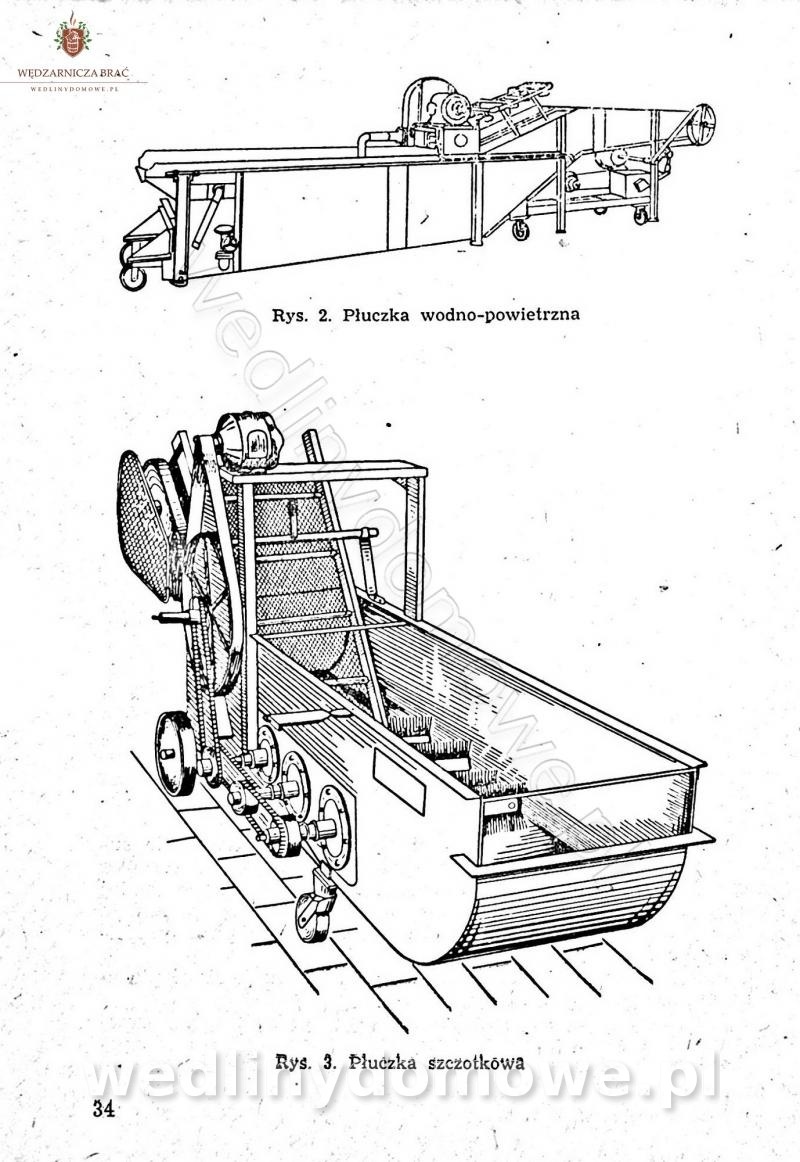
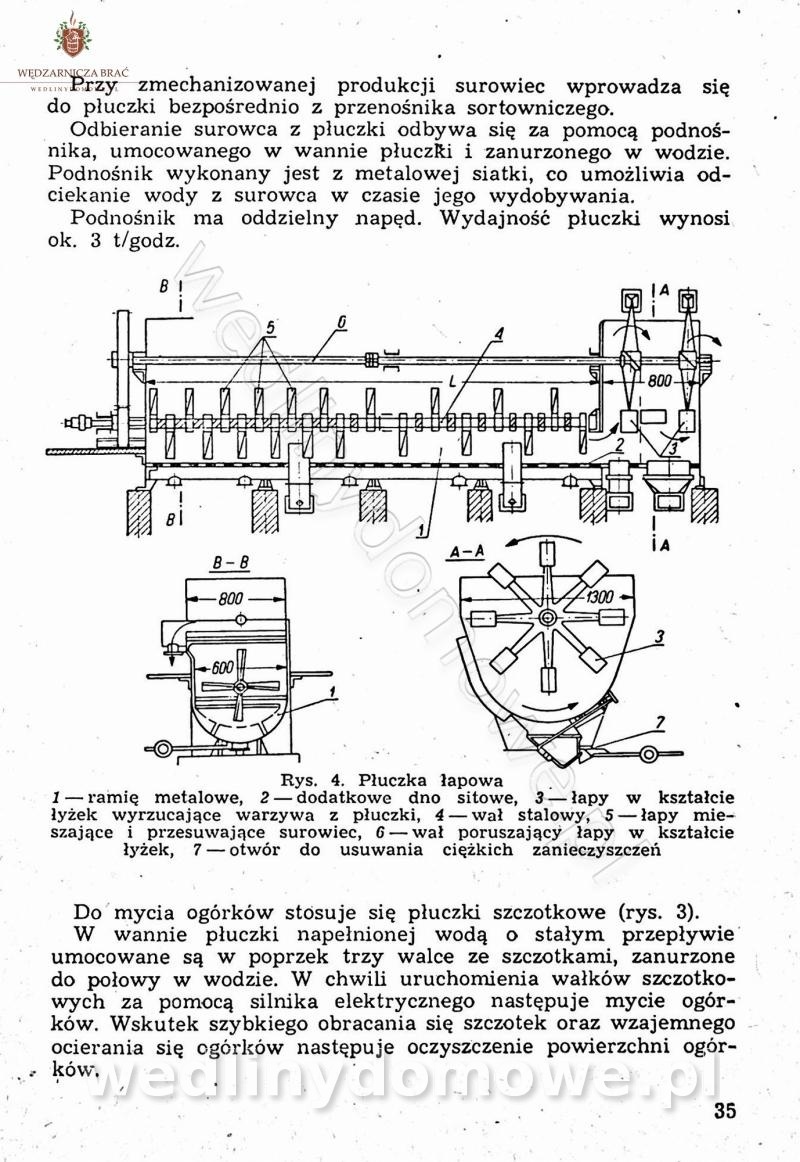








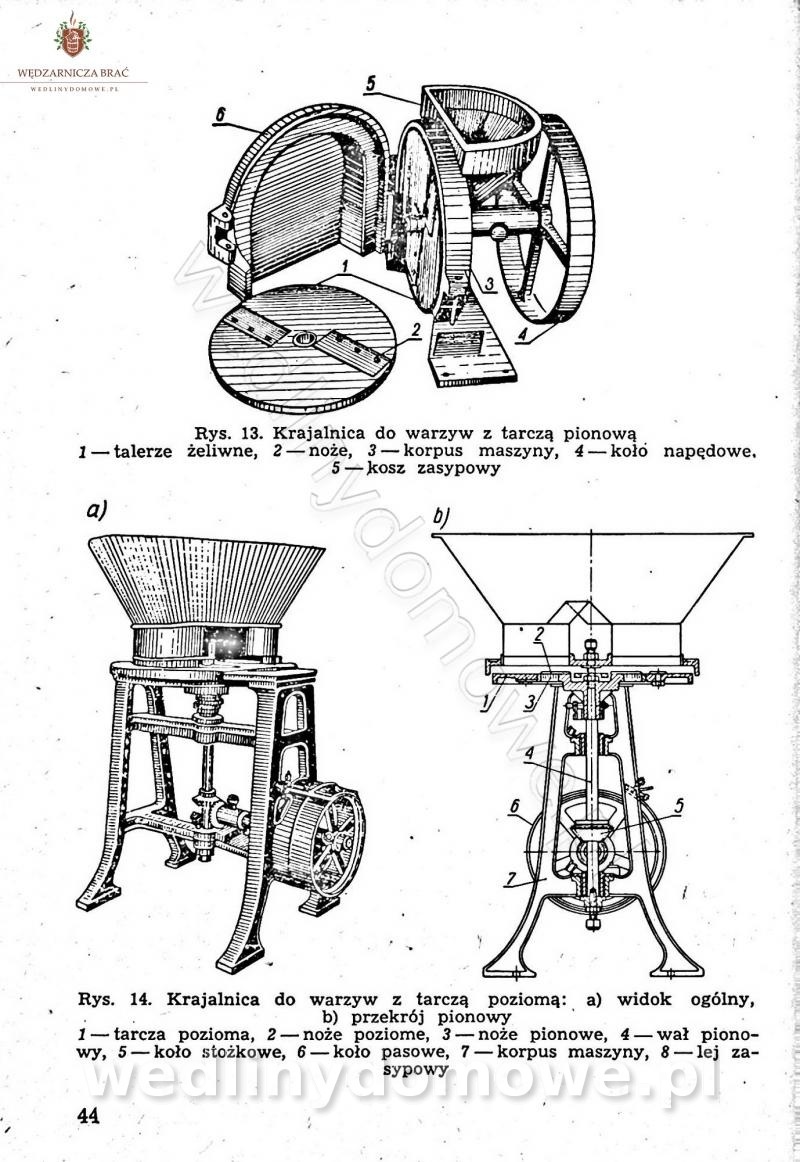

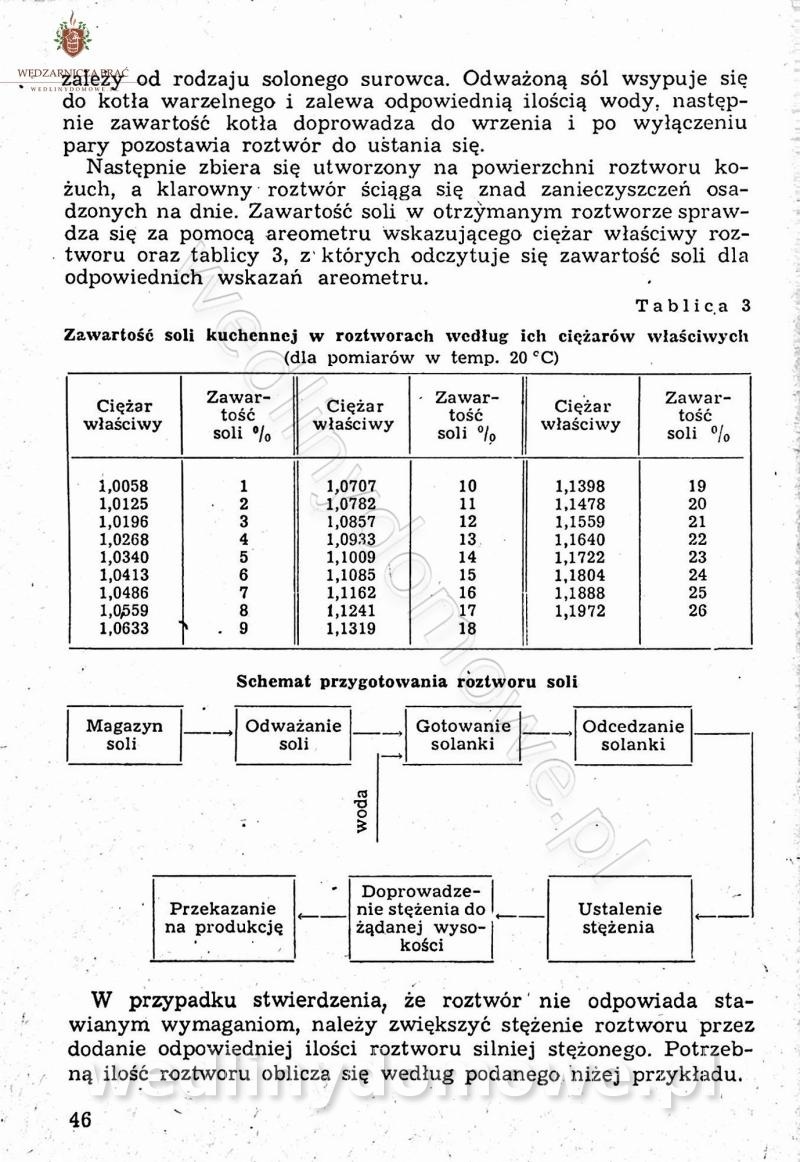



-
Jestem jak najbardziej za. Kto się zajmie tematem?
-
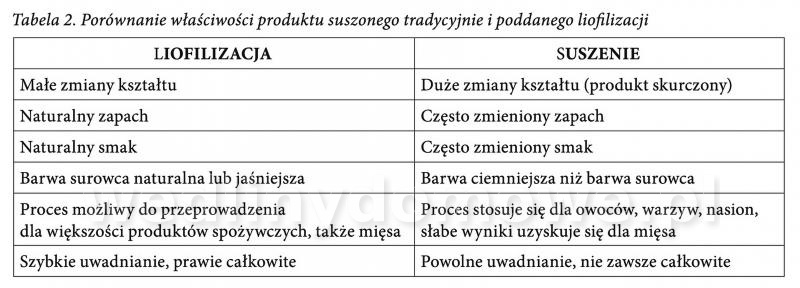
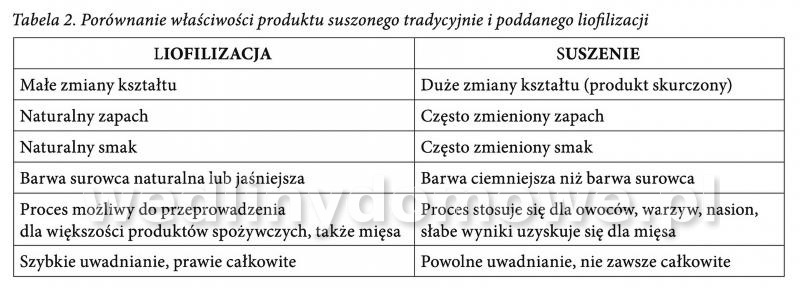
Liofilizacja w przemyśle mięsnym Liofilizowana szynka Słowo "liofilizacja" brzmi tajemniczo, jednak jest to tylko i wyłącznie proces suszenia poprzez mrożenie. Metoda ta, po raz pierwszy została opracowana i zastosowana w latach pięćdziesiątych XX wieku na zlecenie rządu USA zmierzającego do wyprodukowania lekkich wagowo i bogatych we wszystkie niezbędne składniki odżywcze racji żywności dla astronautów i wojska. W Polsce proces ten staje się coraz bardziej popularny. Żywność liofilizowana jest produkowana komercyjnie i wykorzystywana przez ekstremalnych sportowców - podróżników, wspinaczy wysokogórskich i żeglarzy, a także „zwyczajnych” konsumentów. Liofilizacja - co to takiego? Liofilizacja jest jedną z metod suszenia produktu, której głównym założeniem jest usunięcie wody z zamrożonego materiału wskutek procesu sublimacji lodu, z pominięciem fazy ciekłej. Proces ten, zwany także suszeniem sublimacyjnym lub molekularnym, może przebiegać tylko w warunkach ujemnej temperatury oraz przy znacznie obniżonym ciśnieniu (poniżej 200 Pa). Krok po kroku... Suszenie liofilizacyjne żywności obejmuje trzy główne etapy. W pierwszym z nich, wstępnie przygotowany surowiec zostaje poddany zamrożeniu poprzez samozamrożenie, jako skutek obniżenia ciśnienia i wyparowania części wody z żywności kosztem ciepła z niej pobranego. Proces mrożenia można prowadzić w popularnych zamrażarkach. Należy jednak pamiętać, że wybór urządzenia do zamrażania zależy od rodzaju żywności. Produkty w małych kawałkach zamrażane są szybko, w celu utworzenia się małych kawałków lodu, które nie spowodują zniszczenia struktury komórkowej żywności. Natomiast w przypadku żywności płynnej zamrażanie prowadzi się tak, aby powstały skupienia kryształów i kanaliki, ułatwiające ruch pary wodnej. Temperatura mrożenia wynosi od -20 do -40°C i powoduje zabicie większości bakterii chorobotwórczych oraz pleśni. W drugim etapie dokonuje się, tak zwanej próżniowej sublimacji lodu. Zamrożona żywność zostaje umieszczona w komorze niskich ciśnień, na półkach ogrzewanych od wewnątrz, ciśnienie pary wodnej nad powierzchnią żywności redukuje się (poniżej 6-10 Pa), jednocześnie ogrzewając produkt. W tych warunkach kryształki lodu znajdujące się w żywności, sublimują, czyli zamieniają się w parę wodną bez topnienia. Para wodna jest usuwana z komory w sposób ciągły za pomocą skraplacza i pompy próżniowej (suszarka sublimacyjna stanowi połączenie suszenia sublimacyjnego z suszarką próżniową). Suszenie sublimacyjne umożliwia usunięcie z artykułów spożywczych 80-95% wody. W ostatnim, trzecim etapie następuje dosuszanie materiału do żądanej wilgotności końcowej (zawartość wody powinna wynosić około 1-2%), w sposób podobny do suszenia w suszarce próżniowej, w wyniku podwyższenia temperatury żywności do 40-50°C, z utrzymaniem niskiego ciśnienia w komorze. Szybkość liofilizacji zależy od wielu czynników: • grubości warstwy materiału (suchego i zamrożonego), • przewodnictwa cieplnego materiału, • temperatury warstwy wysuszonej (uwarunkowanej termostabilnością materiału), • temperatury frontu sublimacji (zalecana temperatura powinna wynosić od -15 do -45°C), • temperatury kondensatora, • sposobu zamrożenia surowca, • ciśnienia podczas liofilizacji. Liofilizacja mięsa Liofilizacji można poddać każdy produkt i surowiec spożywczy, o ile zawiera on w składzie wodę i składniki zaliczane do tak zwanej suchej substancji, czyli białka, węglowodany i tłuszcze. Najczęściej znaleźć można w sklepach liofilizowane owoce i warzywa, grzyby, sery, jogurty czy ryby. Coraz częściej spotyka się również liofilizowane mięso. W przypadku mięsa poddaje się go kontaktowemu zamrożeniu i wymrożoną wodę (lód) natychmiast sublimuje się pod zmniejszonym ciśnieniem doprowadzając kontaktowo (przy użyciu promienników podczerwieni lub urządzeń mikrofalowych), potrzebną ilość energii cieplnej w celu zrównoważenia jej ubytku spowodowanego sublimacją. W większości stosowanych urządzeń, wszystkie te procesy przebiegają w tej samej komorze, przy czym płyty przewodzące czynnik zamrażający mięso przewodzą kolejno czynnik dostarczający ciepła. Kawałki mięsa uzyskane w procesie sublimacji zachowują swoje rozmiary i kształt, są silnie porowate, zawierają około 2% wody i jednocześnie łatwo ją przyjmują w objętości zbliżonej do utraconej w czasie suszenia. Niestety rozwinięta powierzchnia wysuszonego mięsa, o gąbczastej strukturze, jest bardzo podatna na utlenianie po ekspozycji na powietrzu. Można jednak tego uniknąć wprowadzając do komory w końcowej fazie suszenia gaz obojętny, np. azot i w tych warunkach pakować wysuszone mięso do hermetycznych opakowań. Aby uzyskać granulat mięsny z drobno zmielonego i gotowanego mięsa stosuje się suszenie w przeciwprądzie gorącego powietrza. Taki produkt, zamknięty w szczelnych, odpowietrzonych puszkach jest bardzo trwały i może być wykorzystywany do szybkiego przygotowania wielu potraw. Na rysunku 1 i 2 przedstawiono liofilizaty mięsne. Dania mięsne liofilizowane najczęściej występują w postaci gotowych dań (jedno- lub dwuosobowych porcji). Posiłki takie, wyróżnia niska masa (nie więcej niż 150-200 g) i łatwość w transporcie (grube foliowe bądź mylarowe woreczki). Liofilizaty mięsne uważane są za pełnowartościowe posiłki, które mogą być wykorzystywane jako część zdrowej i odpowiednio zbilansowanej diety, dlatego też są chętnie wybierane przez osoby aktywne fizycznie i dużo podróżujące. Zalety i wady liofilizatów mięsnych Produkty mięsne suszone metodą sublimacji zachowują swoje pierwotne właściwości organoleptyczne (smak, zapach, barwę, aromat) oraz pełną wartość odżywczą i biologiczną (enzymy, aminokwasy, witaminy, minerały chelatowane). W porównaniu z tradycyjnym suszeniem (tabela 2) preparaty zliofilizowane cechują się bardzo małą zawartością wody (1- 2%), co zabezpiecza je przed niepożądanym rozwojem różnych rodzajów mikroflory. Do żywności otrzymanej w ten sposób nie dodaje się środków konserwujących, co ma istotne znaczenie przy wytwarzaniu "zdrowej" żywności. Poprzez porowatą strukturę i niską zawartość wody są bezpieczne mikrobiologicznie i zachowują praktycznie pełną rehydratyzację. Dzięki dużej powierzchni wewnętrznej, odznaczają się również bardzo dobrą rozpuszczalnością. Często też są silnie higroskopijne co oznacza, że są lepiej wchłanianie przez organizm, niż inne produkty konserwowane tradycyjnymi metodami, np. poprzez pasteryzację czy sterylizację (wyniszczającą naturalne formy enzymów, białek roślinnych i chelatów wiązanych z minerałami). Dodatkowo, dzięki liofilizacji okres przechowywania produktów mięsnych zapakowanych w szczelne opakowanie (w każdych warunkach klimatycznych) może wynosić nawet kilka lat. Niestety za wadę produktów liofilizowanych uznać należy fakt, iż koszt ich wyprodukowania jest dość wysoki, co związane jest z koniecznością zastosowania specjalistycznego sprzętu. Proces suszenia sublimacyjnego cechuje się też znaczną energochłonnością. Kolejnym mankamentem jest to, że mięsa które zawierają dużą ilość tłuszczów, są podatne na zmiany oksydacyjne. Z tego powodu do liofilizacji, należy wybierać tylko chude mięsa. Dodatkowo porowata struktura produktów liofilizowanych stanowi duży problem w trakcie przechowywania, ponieważ zwiększa powierzchnię kontaktu fazy stałej z tlenem. Pory wypełnione są powietrzem, a tlen łatwo przenika do wnętrza struktur, przez co oksydacja tłuszczu zachodzi w całej objętości. Co więcej, zarówno pierwotne (nadtlenki), jak i wtórne (głównie aldehydy) produkty utleniania tłuszczu są bardzo reaktywne i łatwo wchodzą w reakcje z niemal wszystkimi składnikami żywności. Dotyczy to zwłaszcza białek, które decydują o prawidłowym funkcjonowaniu naszego organizmu. Zachowanie ich wysokiej wartości odżywczej jest konieczne podczas produkcji przetworów mięsnych, ponieważ białka mięsa są pełnowartościowe i zawierają niezbędne aminokwasy. Warto jednak dodać, że liofilizacja produktów spożywczych z roku na rok jest udoskonalana. Aby ograniczyć „jełczenie” tłuszczów stosuje się dodatkowe metody ograniczające ten negatywny proces. Obok odpowiednich warunków pakowania (np. w atmosferze modyfikowanej) i materiałów opakowaniowych nieprzepuszczalnych dla tlenu i światła, obecnie najskuteczniejszą metodą jest dodatek związków o działaniu przeciwutleniającym. Z uwagi na to, że zastosowanie syntetycznych przeciwutleniaczy (np. BHT, BHA) w przetwórstwie żywności podlega ścisłym regulacjom prawnym, poszukuje się nowych źródeł związków o aktywności przeciwutleniającej. Polecany jest dodatek antyoksydantów naturalnych - witamin o właściwościach przeciwutleniających (witaminy C i E) oraz związków polifenolowych ekstrahowanych z wielu roślin, w tym ziół i przypraw (rozmaryn, tymianek). Coraz więcej polskich firm zajmujących się produkcją artykułów spożywczych inwestuje w maszyny służące do liofilizacji, których na rynku polskim przybywa. Obecnie liofilizaty stały się też bardziej dostępne - sięgnąć może po nie praktycznie każdy. Ich cena nie przeraża przeciętnego konsumenta, gdyż konkurencja firm na rynku jest coraz większa. Mimo że, jedzenie i preferowane smaki to dość indywidualna sprawa, popularność mięsnych produktów liofilizowanych wciąż wzrasta. Może nadszedł czas na wypróbowanie tych dań we własnej kuchni? Autorki: dr inż. Agnieszka Starek mgr inż. Agata Blicharz-Kania




-
Receptura produkcji 1000 kg gołąbków w sosie pomidorowym: 1. Mięso wieprzowe bez kości - 224 kg 2. Mięso cielęce bez kości - 35 kg 3. Smalec wieprzowy - 6 kg 4. Koncentrat pomidorowy 20% - 70 kg 5. Cukier - 3,5 kg 6. Sól kuchenna biała - 12 kg 7. Mąka pszenna 50% - 14 kg 8. Cebula świeża oczyszczona - 23,5 kg 9. Pieprz naturalny - 0,70 kg 10. Ziele angielskie - 0,20 kg 11. Liście laurowe - 0,05 kg 12. Mięso wołowe bez kości - 48 kg 13. Masło wyborowe - 5 kg 14. Marchew świeża oczyszczona - 4 kg 15. Seler świeży oczyszczony - 3,2 kg 16. Pietruszka świeża oczyszczona - 3,1 kg 17. Ryż - 34 kg 18. Kapusta biała w główkach - 625 kg Doczytałem, że wszystkie dania warzywno mięsne sterylizuje się 10 + 50 + 10 w temperaturze 117 st.C. Szczegółową recepturę trzeba będzie odszukać w dalszej części książki: Poradnik Technologa Przetwórstwa Owocowo-Warzywnego. W wolnej chwili postaram się zeskanować i udostępnić całą książkę liczącą 500 stron.
-
NIE. Proszę sobie ustalać ceny i inne techniczne sprawy na PW. Tutaj wrzucamy jedynie zdjęcia i opis książki.
-
Tutaj chyba nikt nie kupi, gdyż ta książka od kilku lat jest na forum: /topic/14285-wsp%C3%B3lnie-z-cookiem-gotujemy-w-kambuzie/
-
Kotły i menażki. Żołnierskie smaki dawniej i dziś.
Maxell odpowiedział(a) na Maxell temat w Polska tradycja masarska
















































-
Pomijając wszystkie pozostałe fakty związane z Twoim zaangażowaniem w życie forum, to stwierdzam, iż nie chce Ci się nawet poszukać odpowiednich tematów.
-
Kotły i menażki. Żołnierskie smaki dawniej i dziś.
Maxell opublikował(a) temat w Polska tradycja masarska



















































-
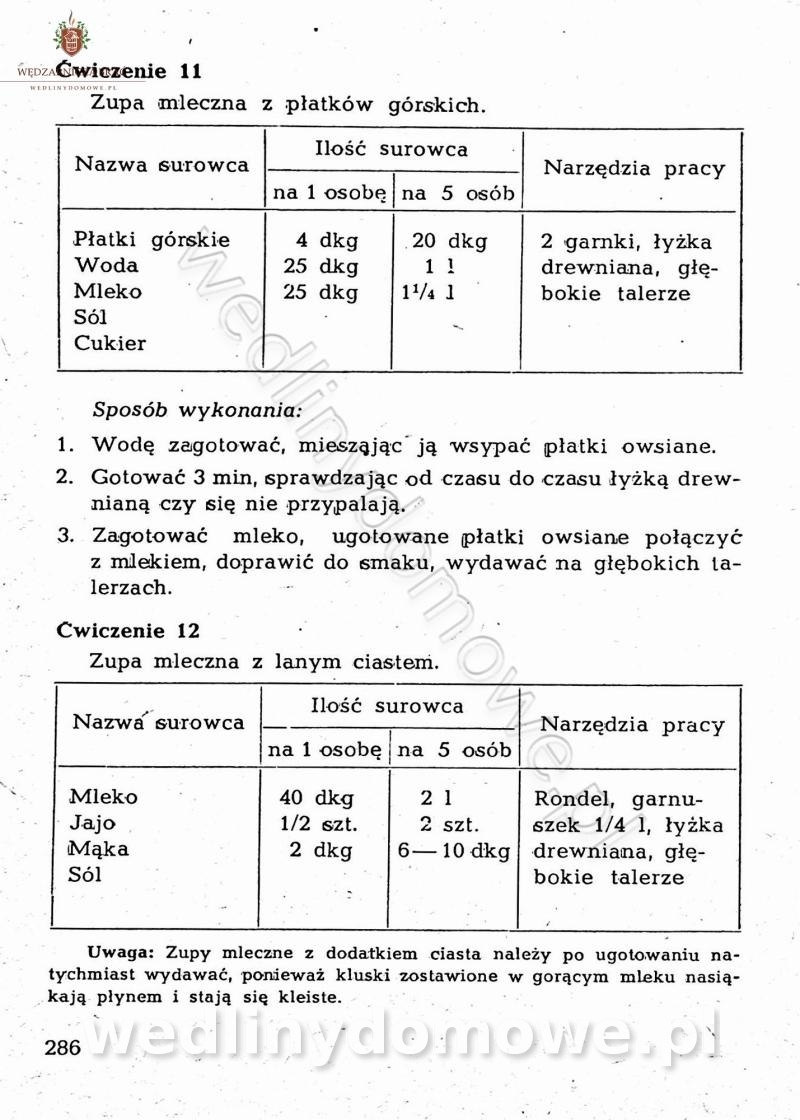
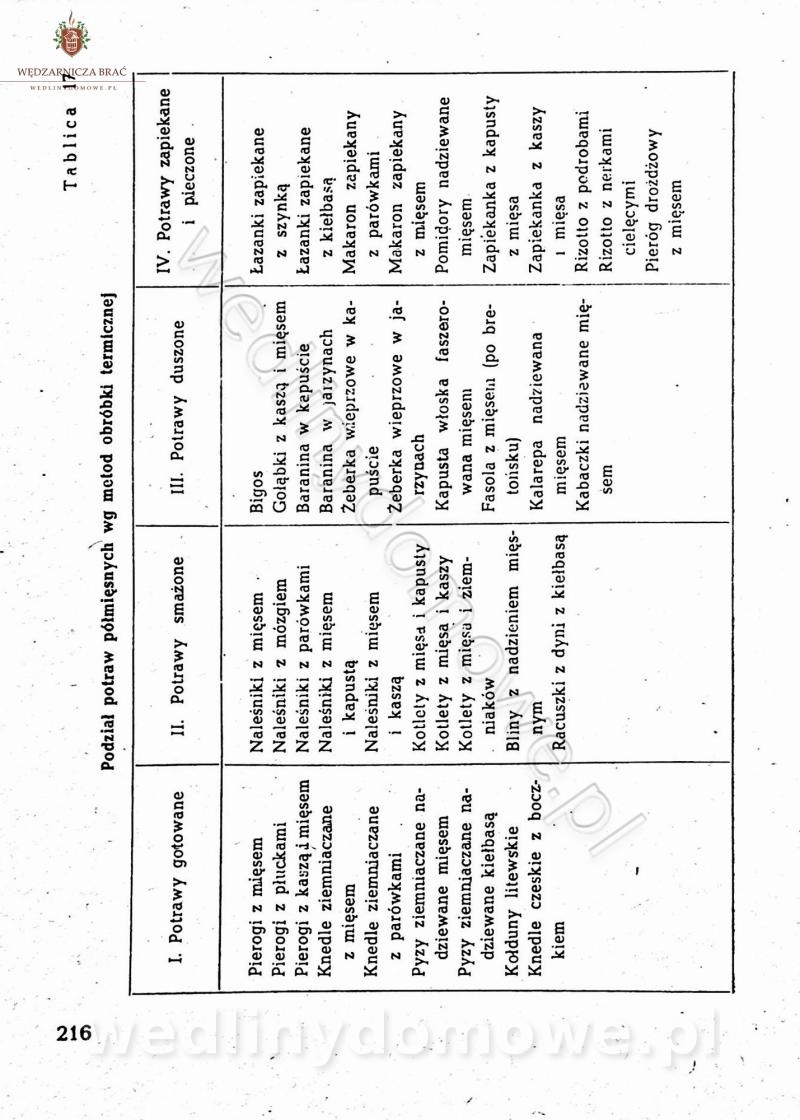
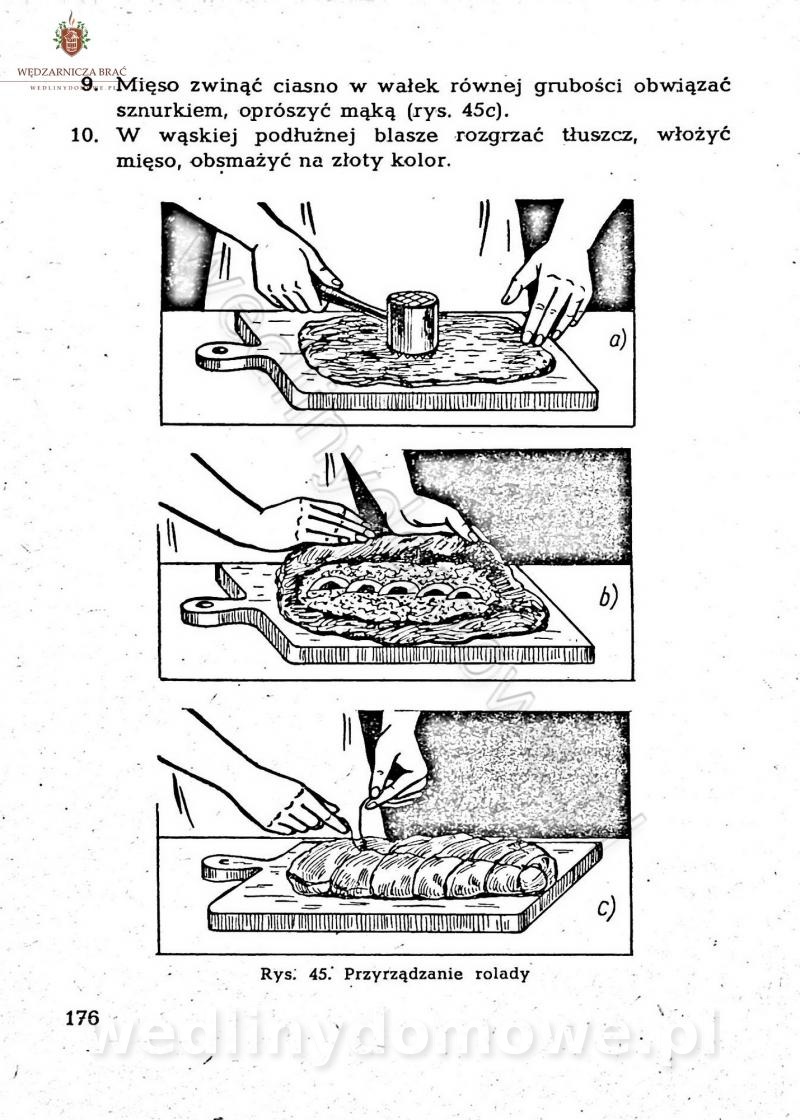
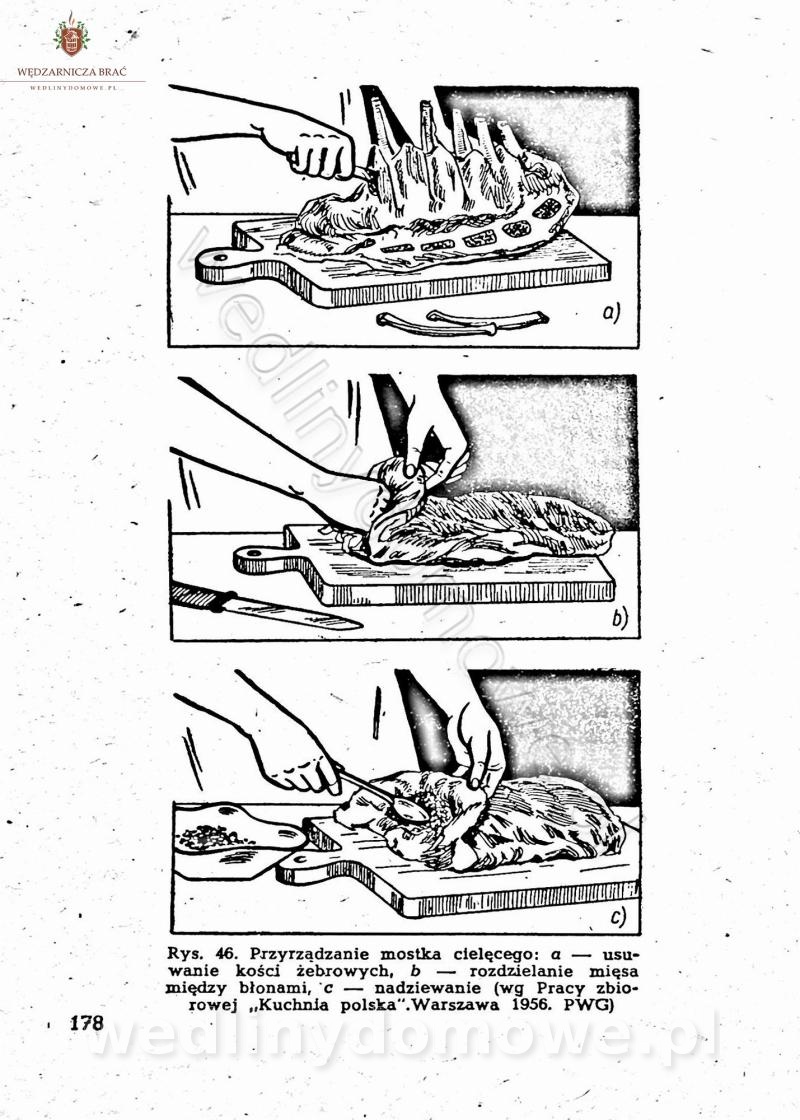
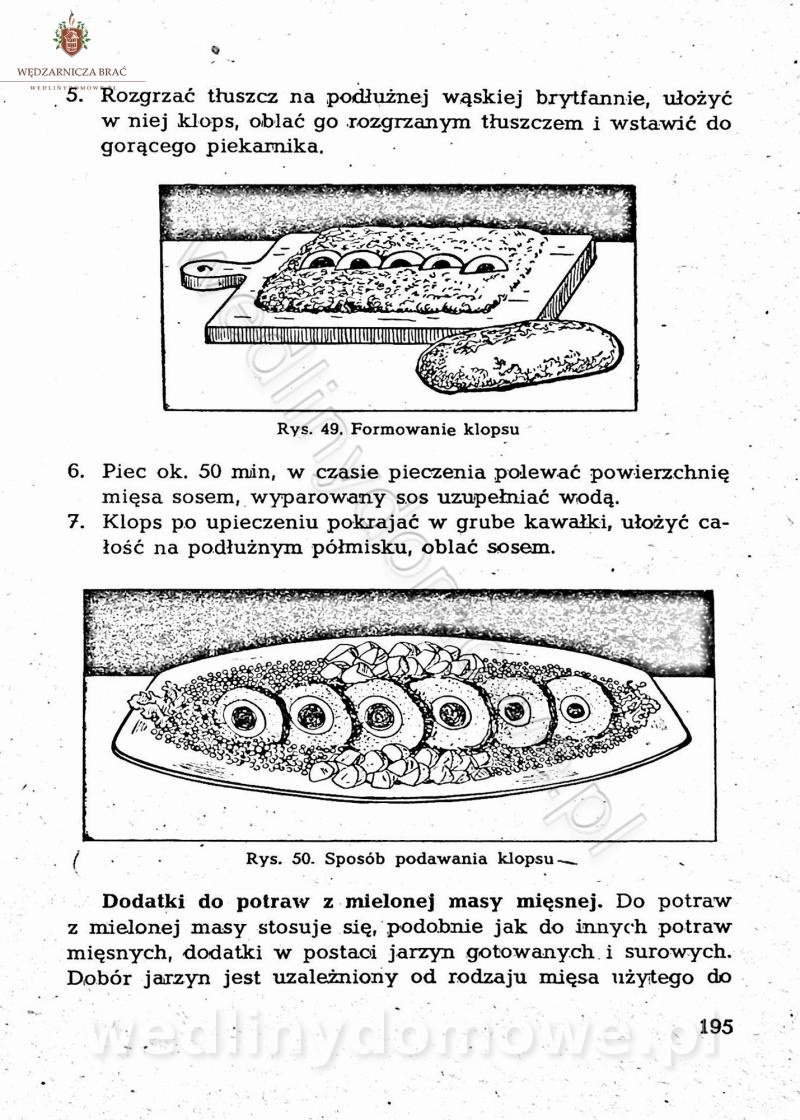
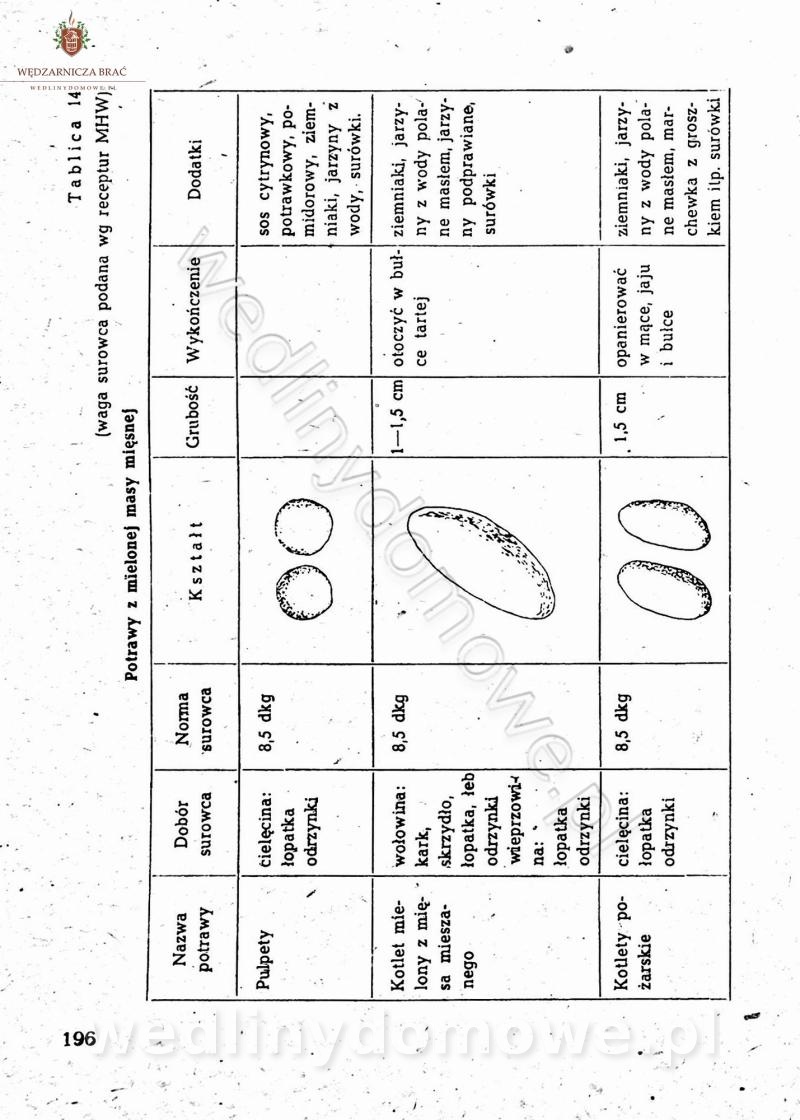
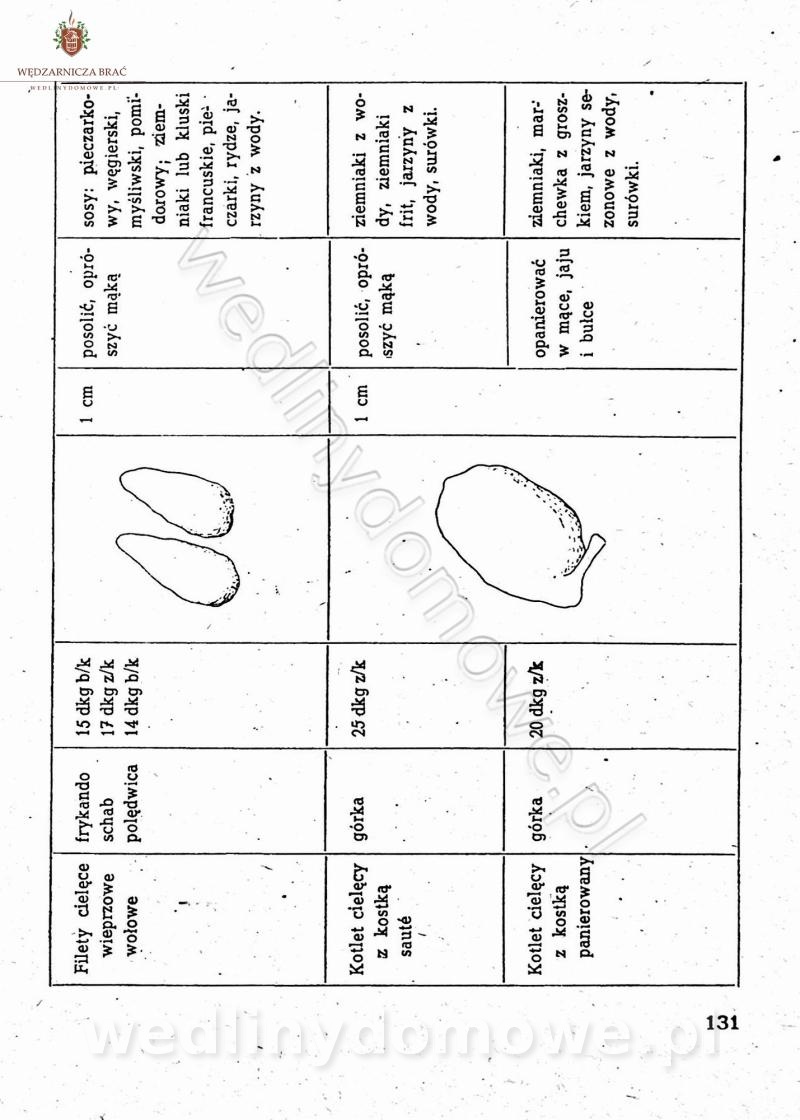
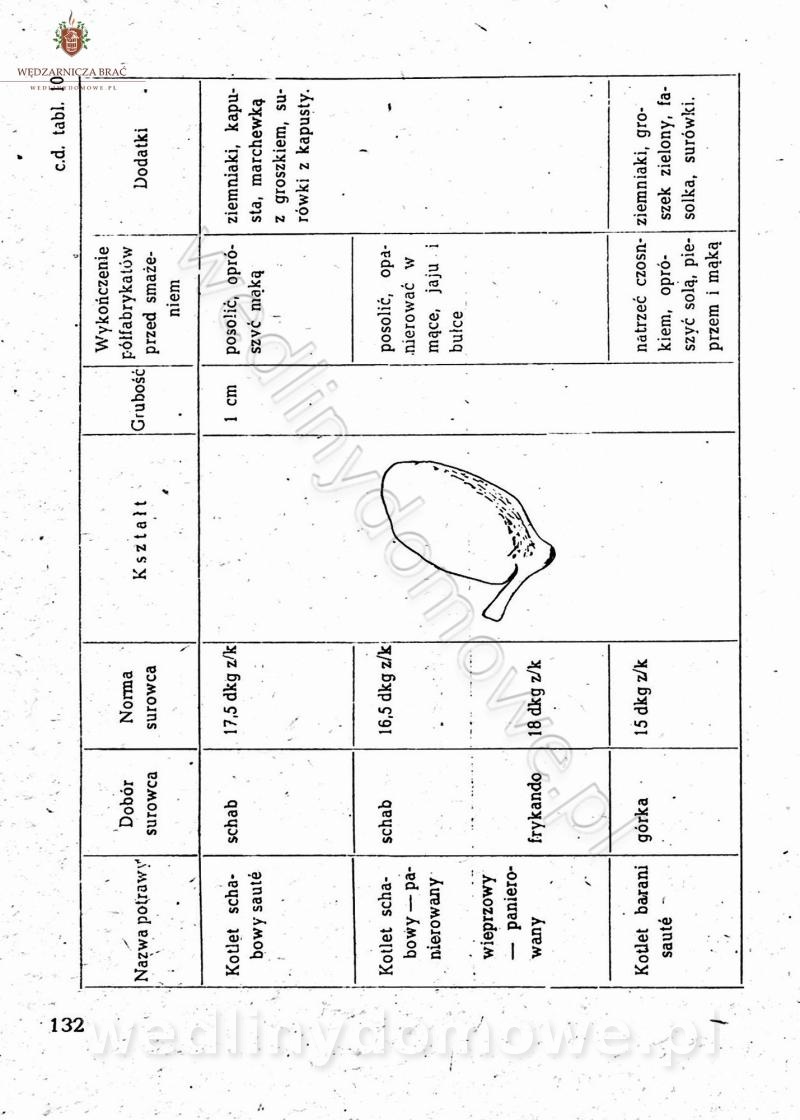
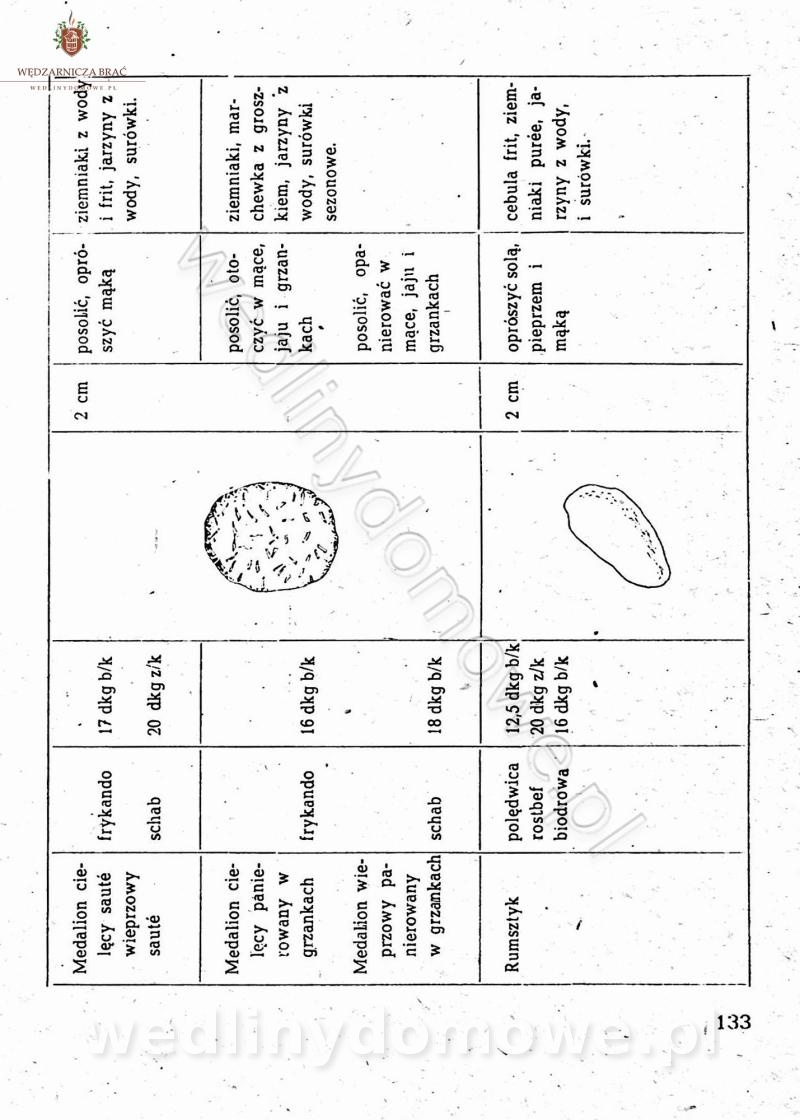
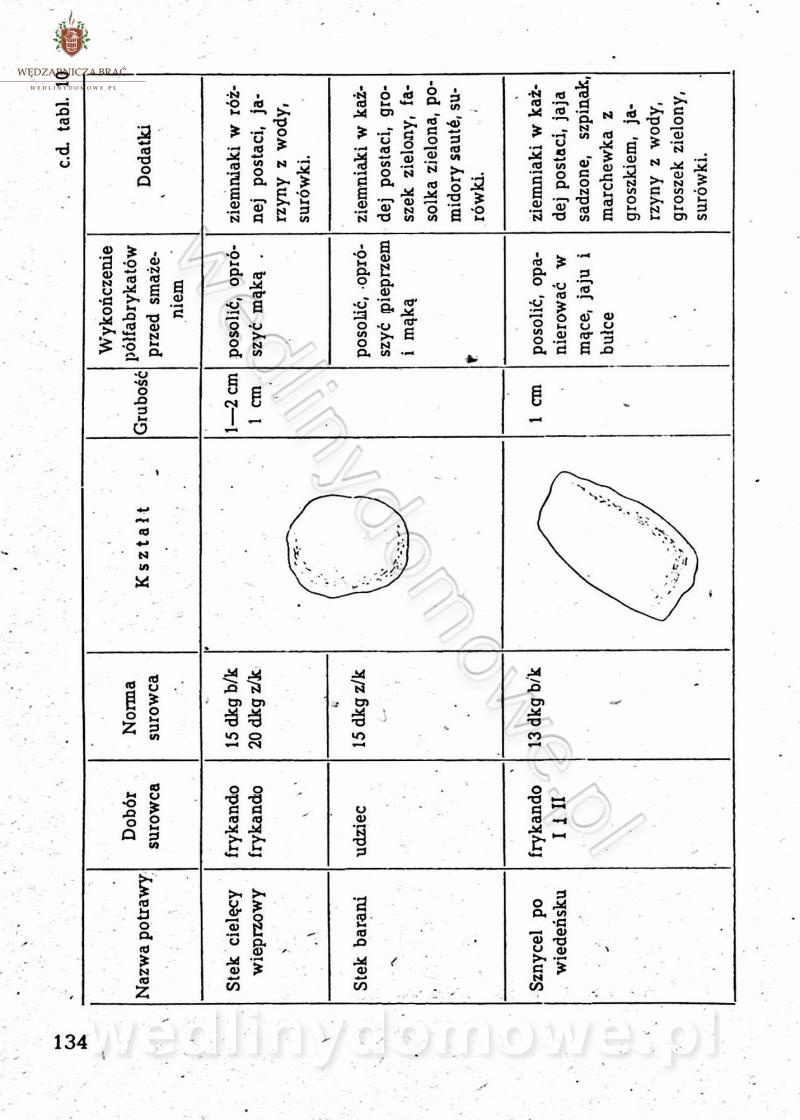
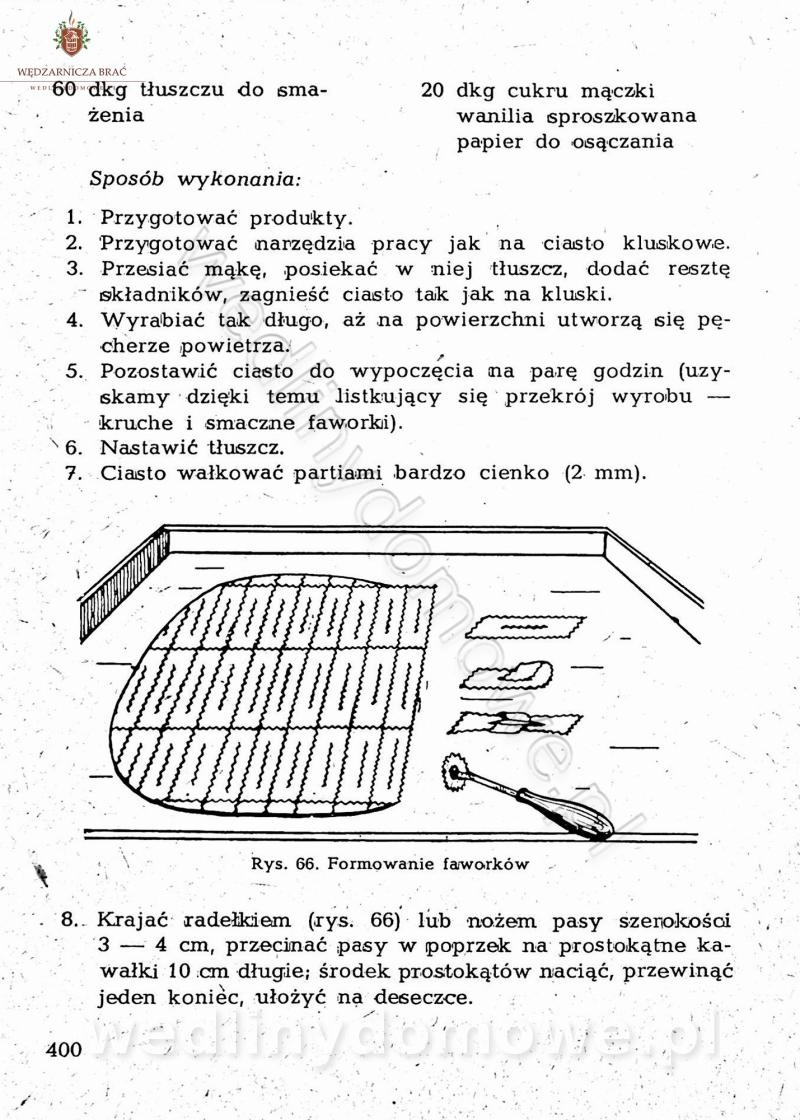
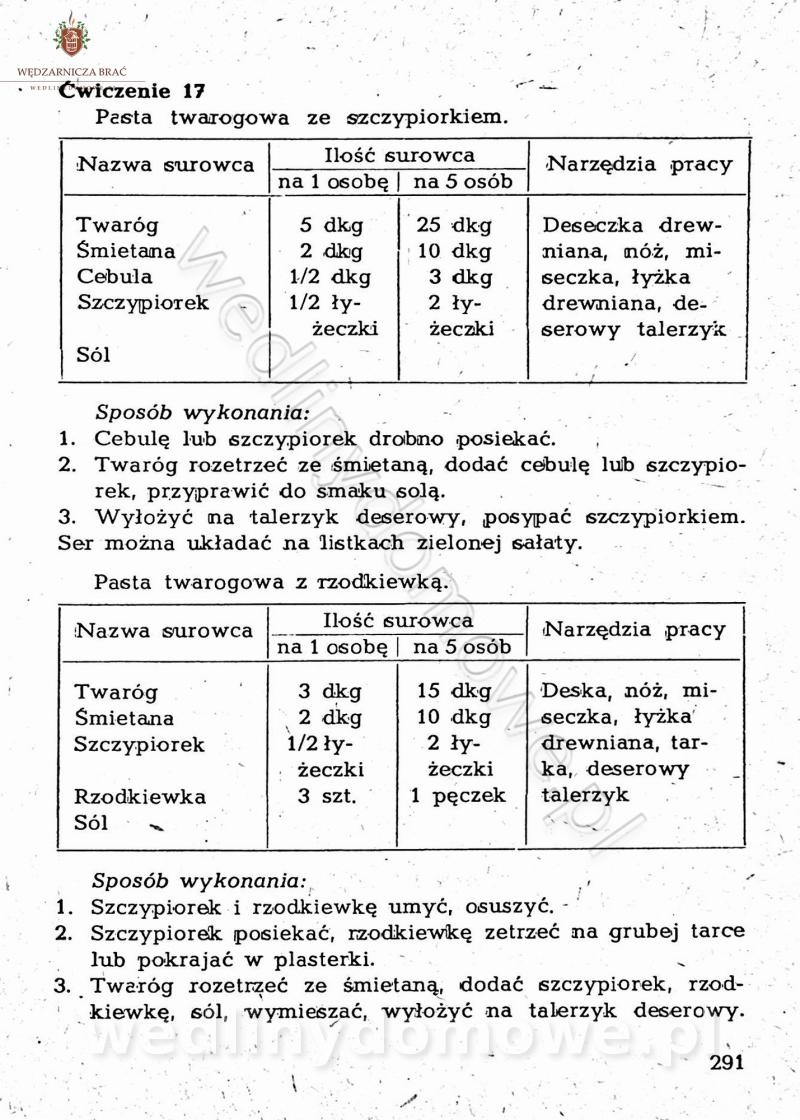
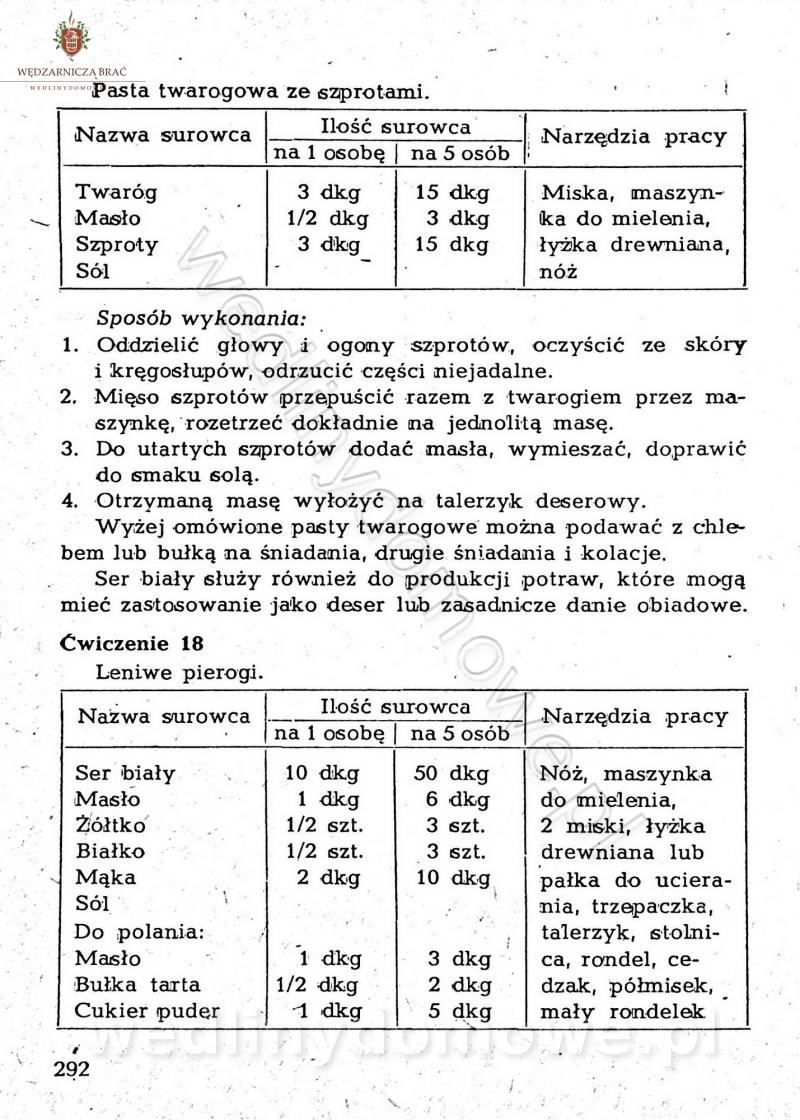
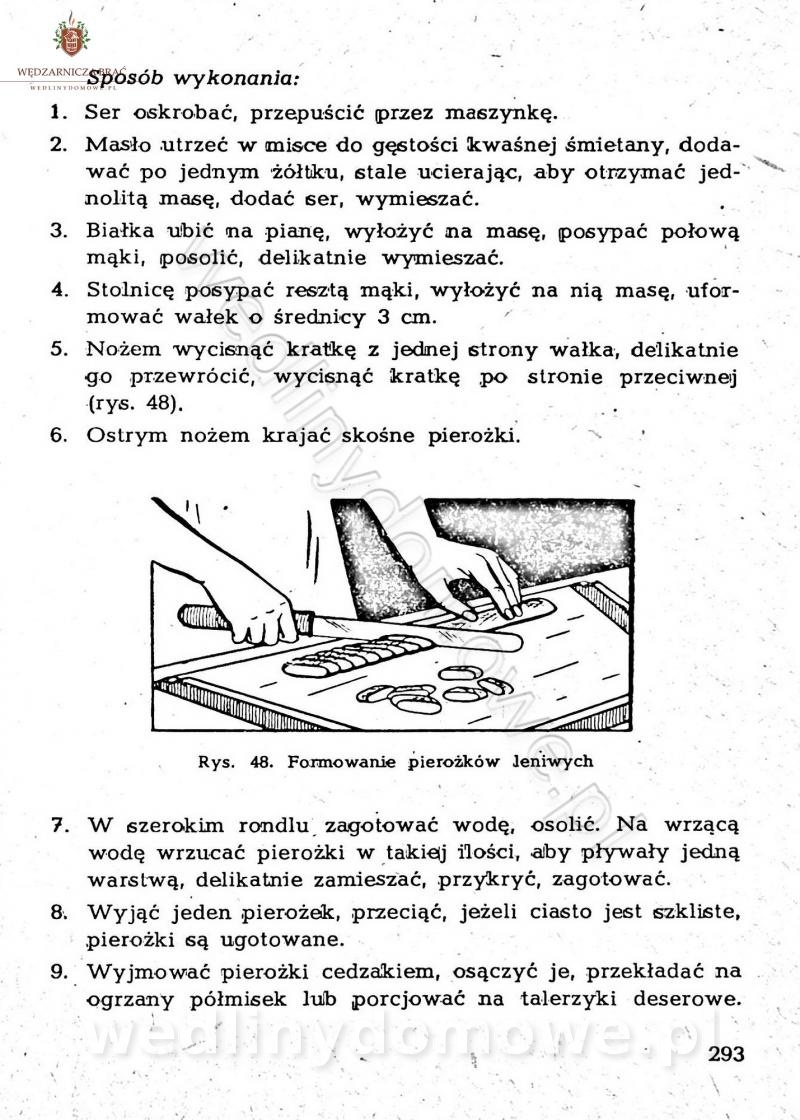
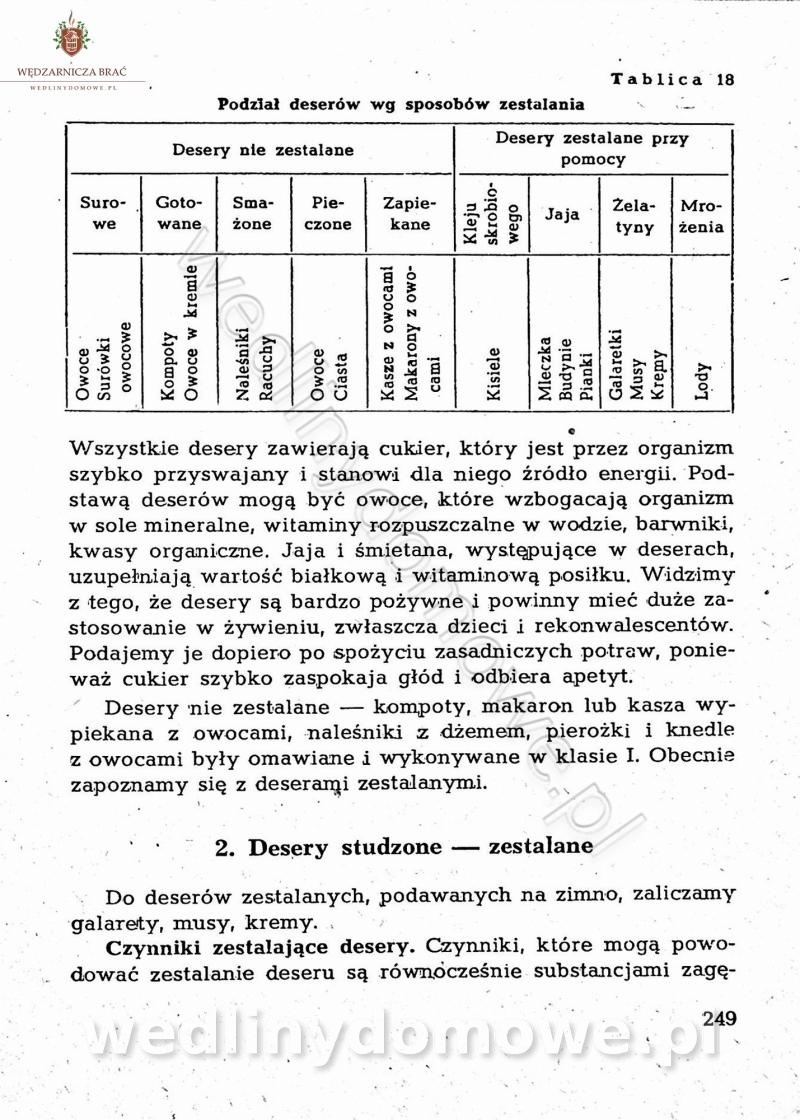
[Rok 1958][Tom 2] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie
Na potrzeby portalu wedlinydomowe.pl opracował Maxell

























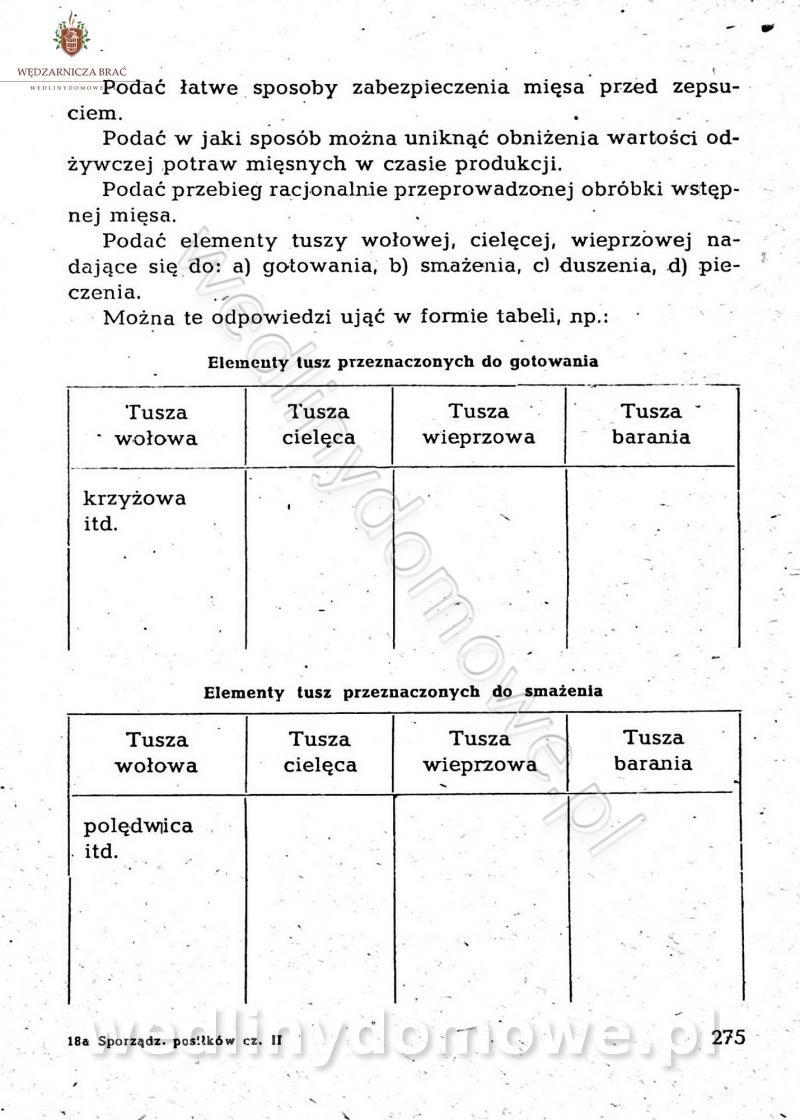











-
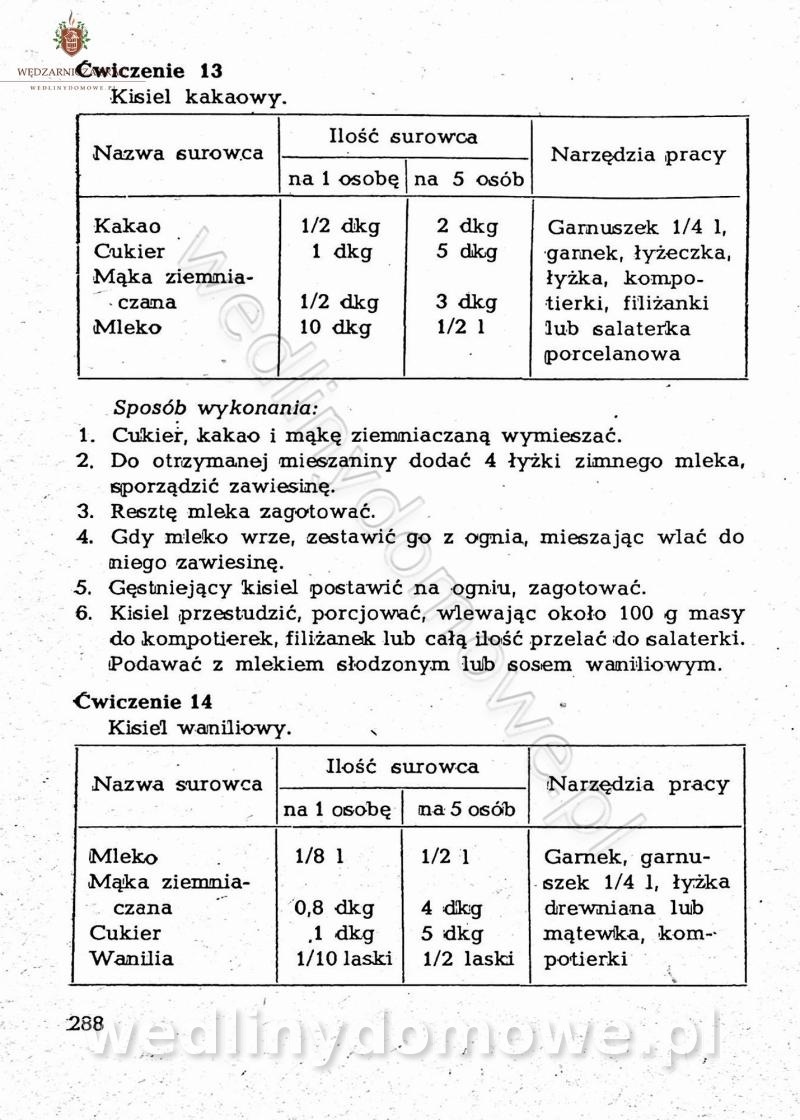
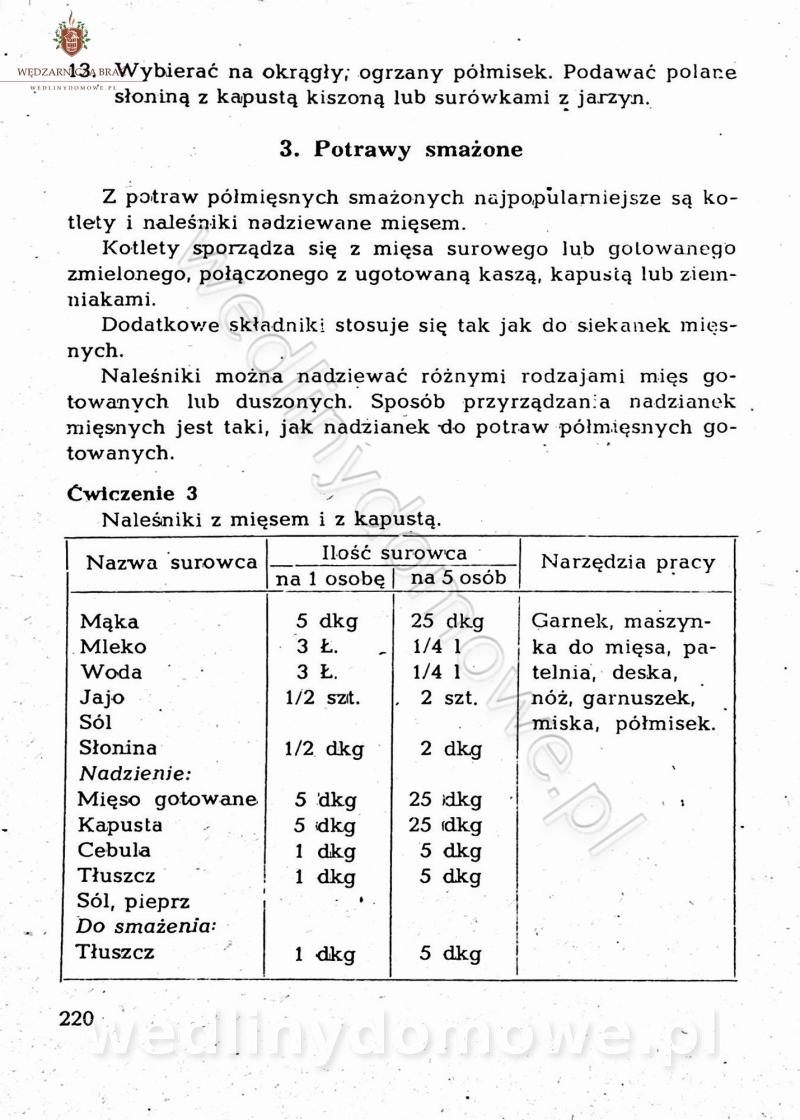
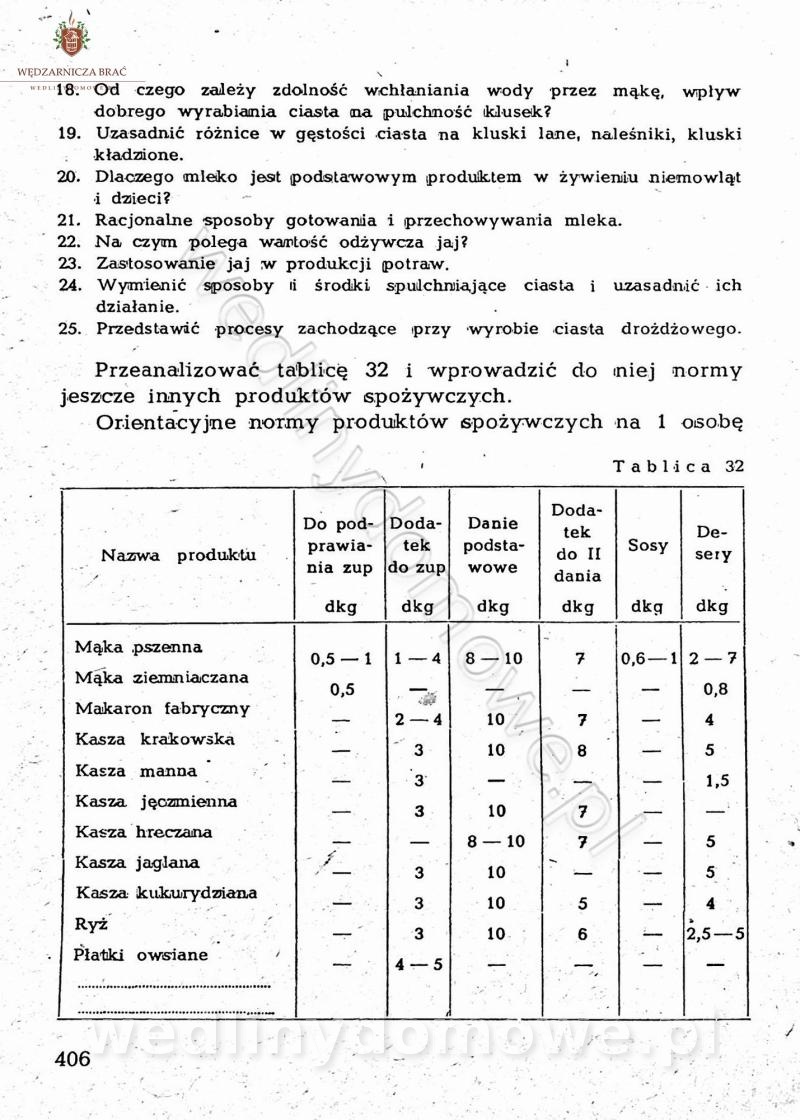
[Rok 1958][Tom 2] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie


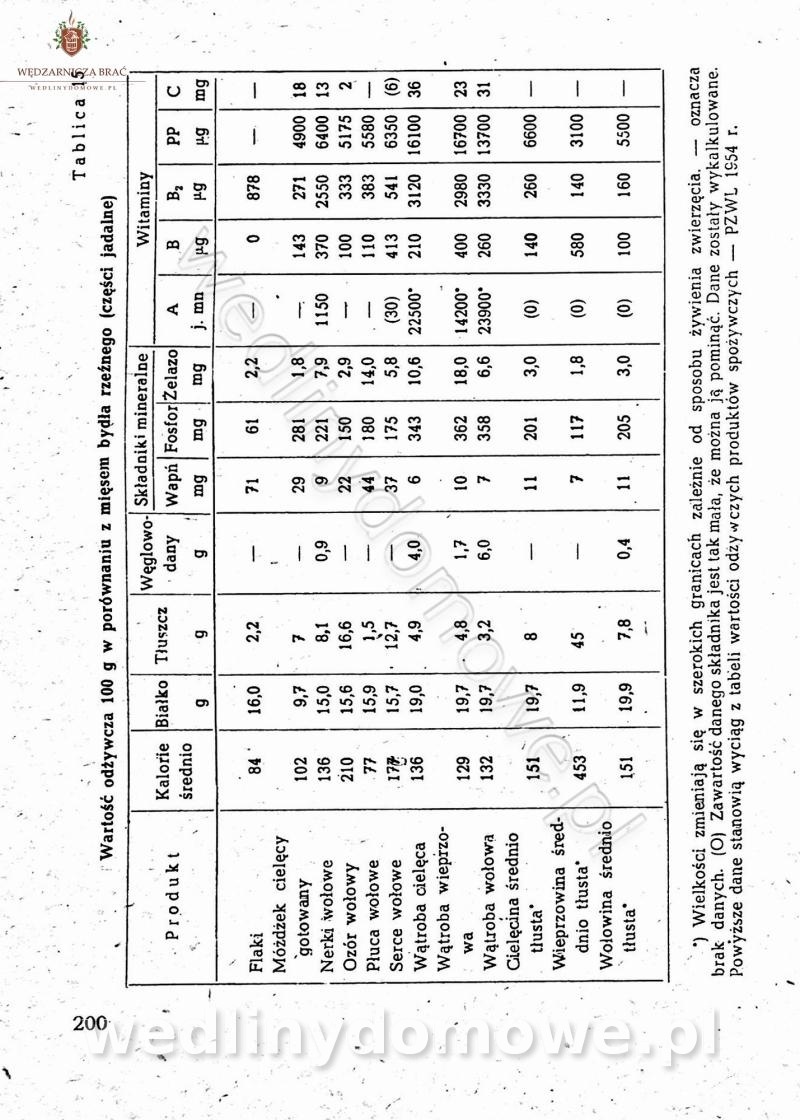












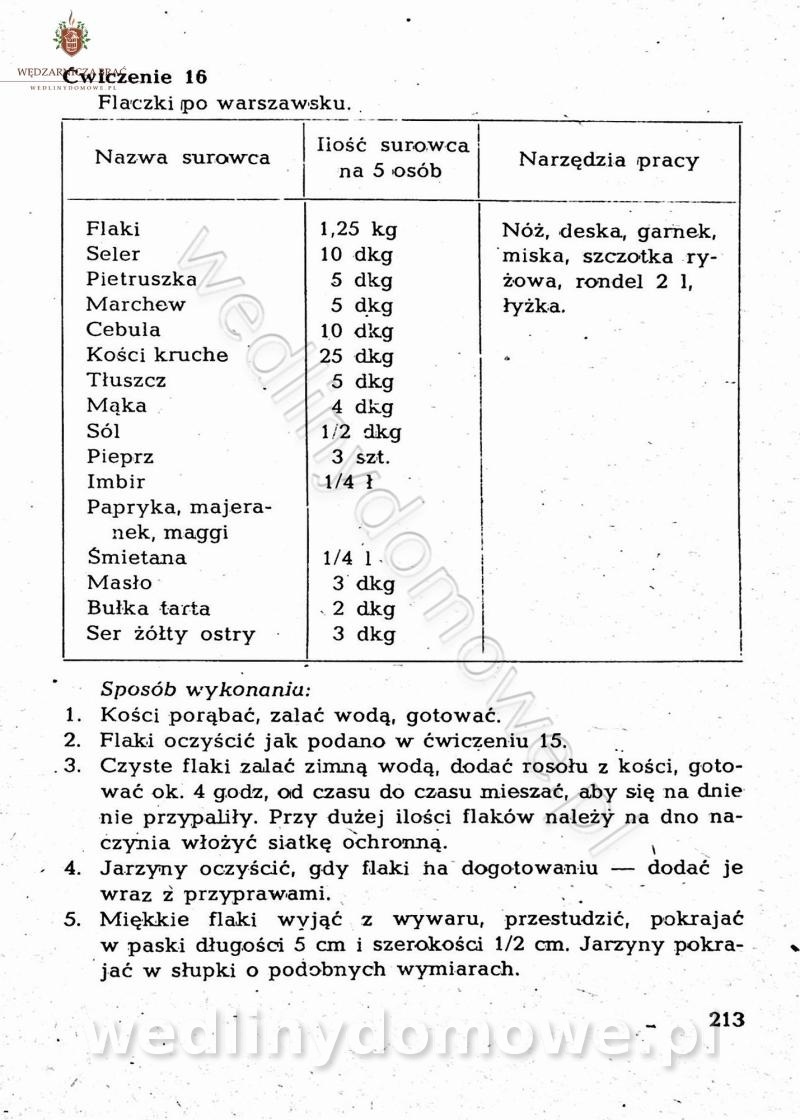


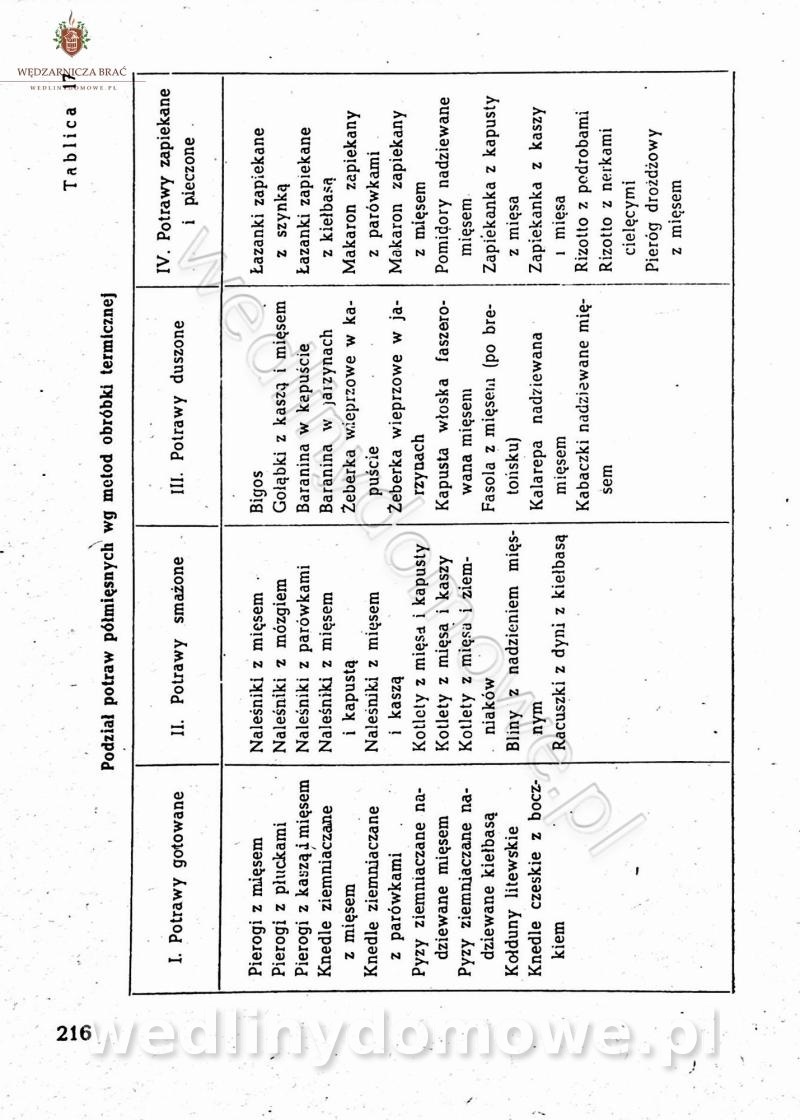



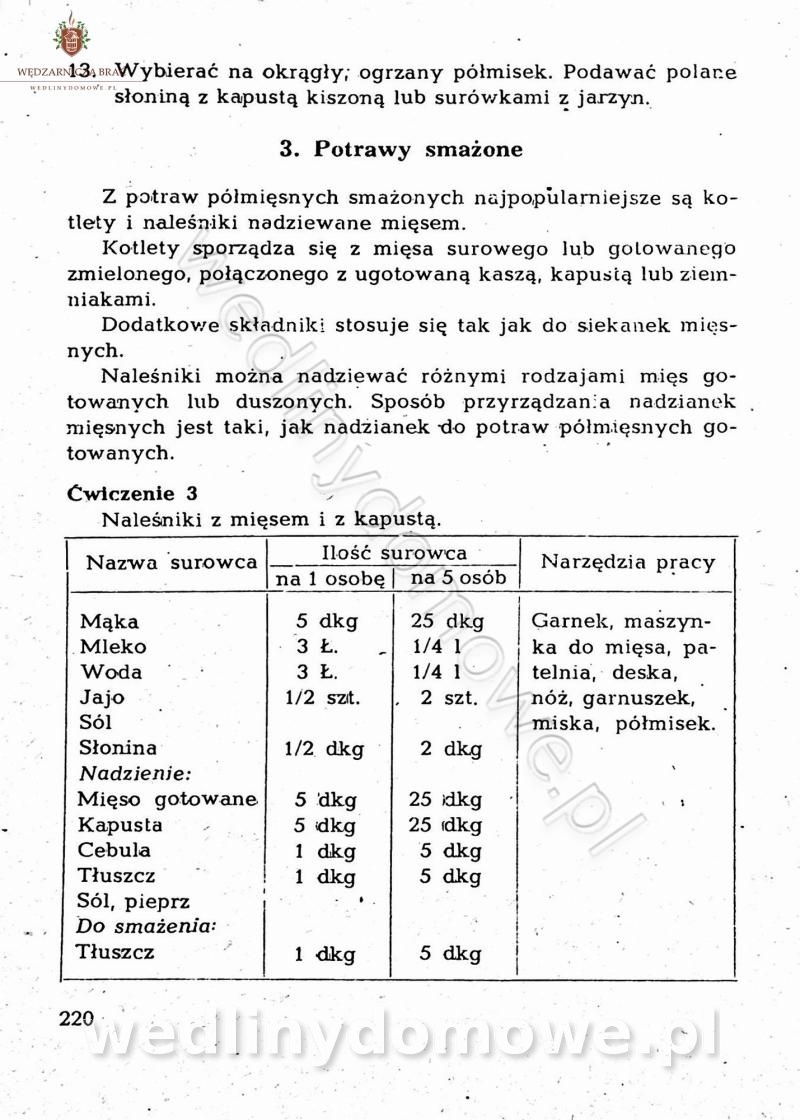





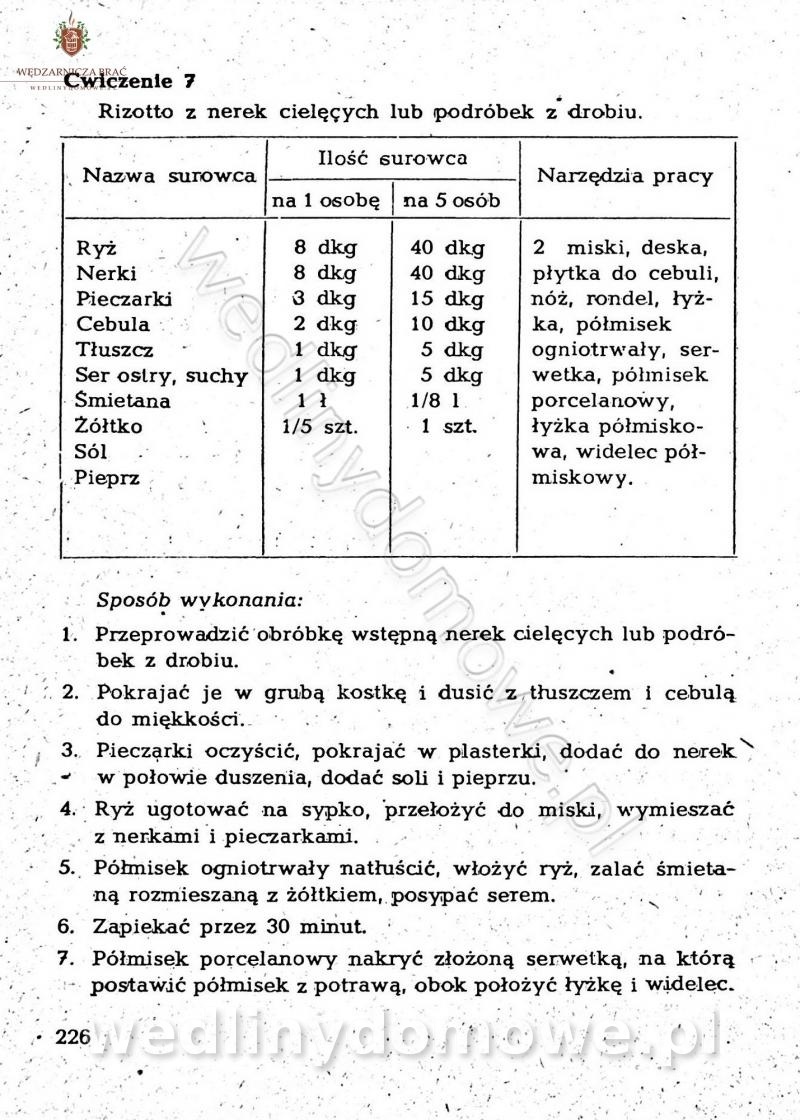
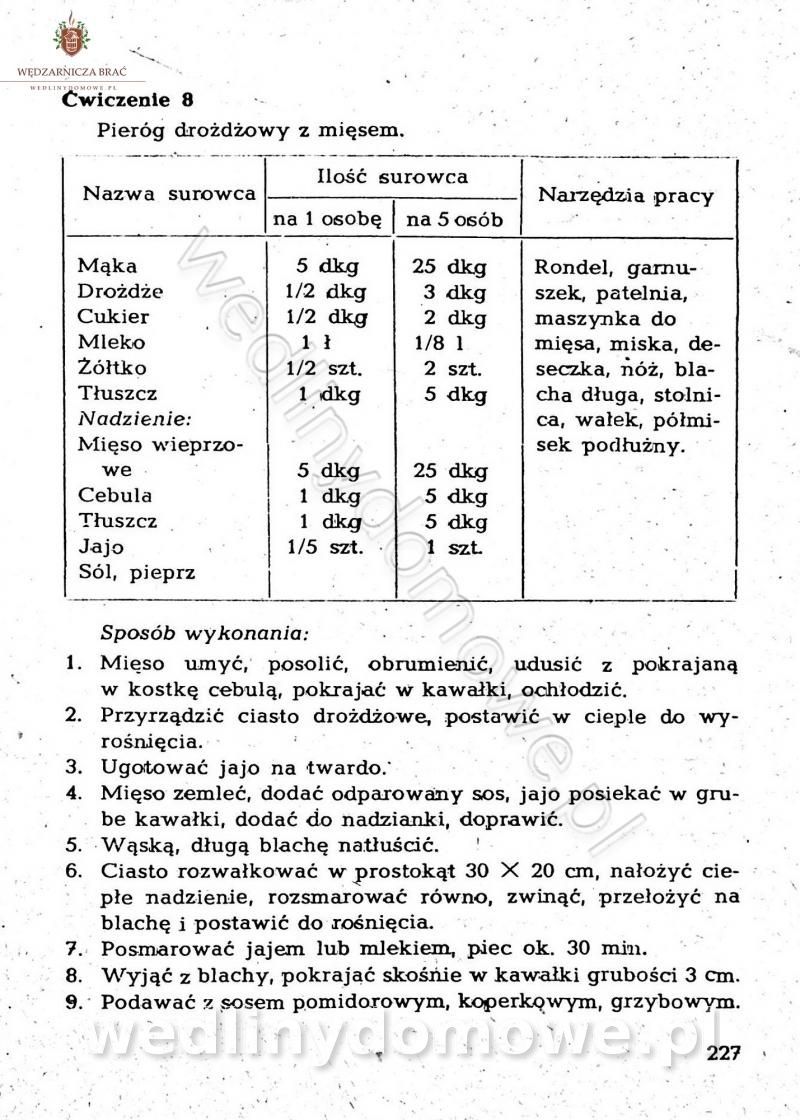


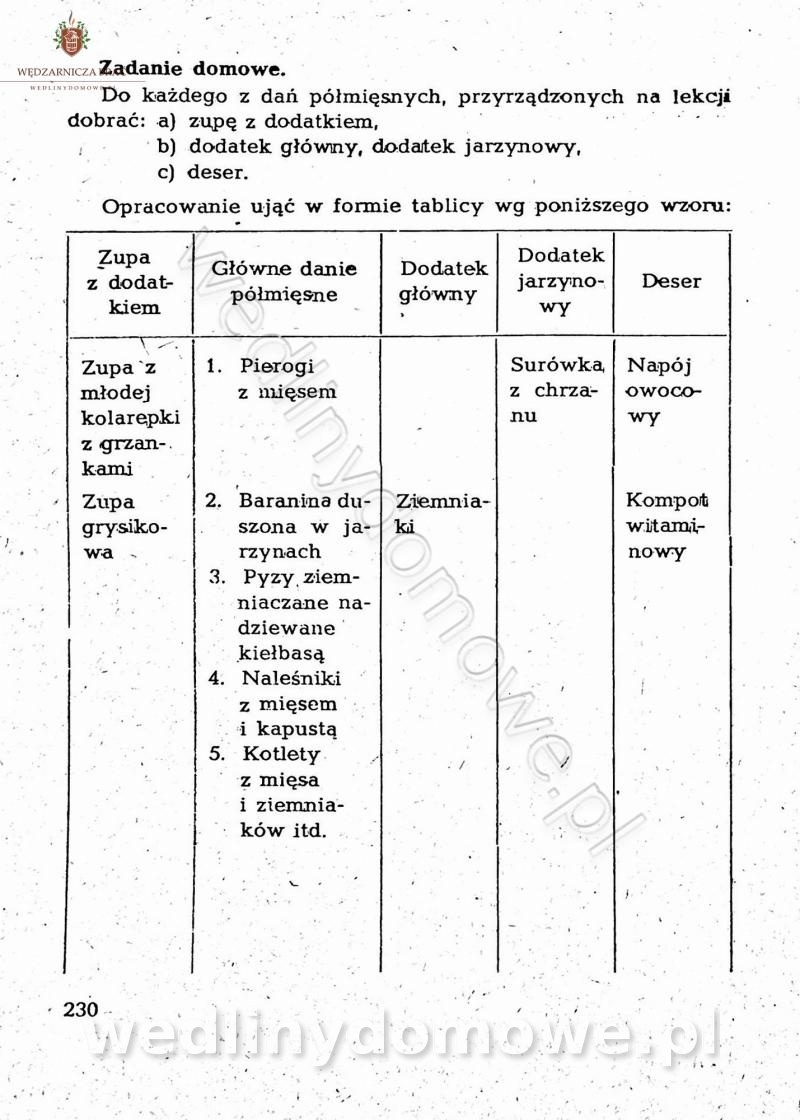



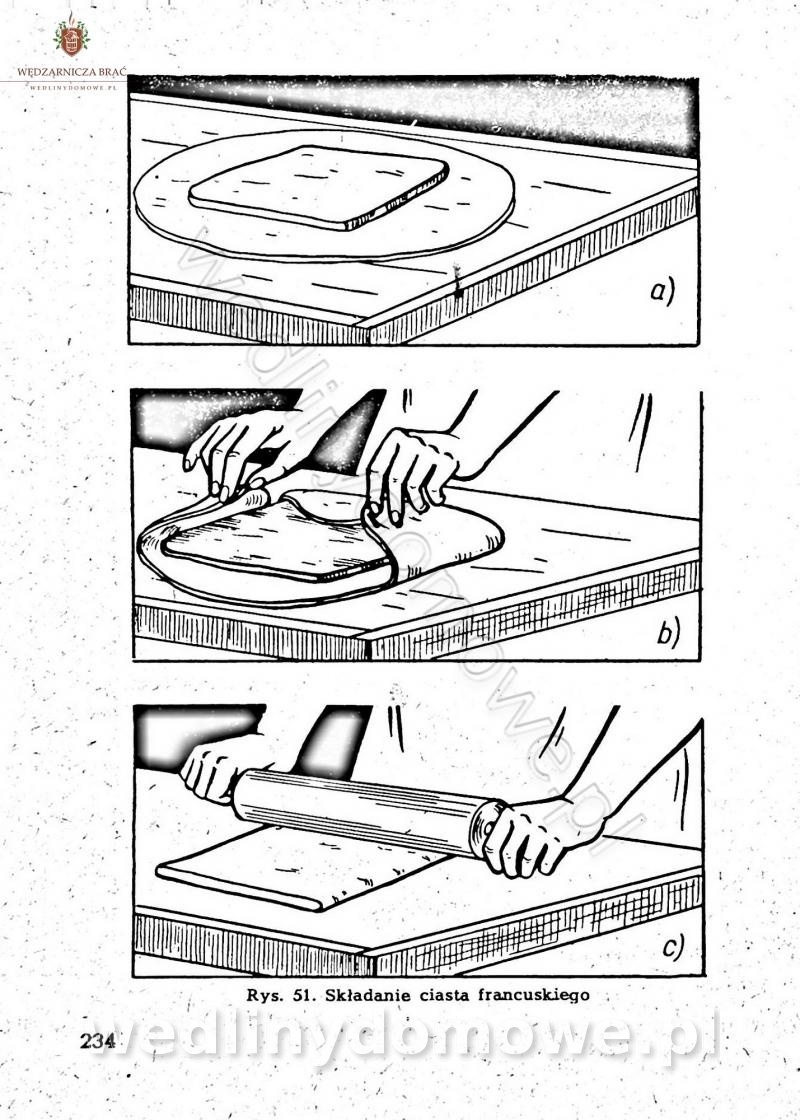












-
Chce się Wam jeszcze brnąć w temat? Już powolutku wracamy na nasze tory i zamiast tematu dymogeneratora, zaczynamy sami się obrażać. Zobaczymy, czy Kolega będzie miał na tyle cywilnej odwagi, by wejść na forum i przeprosić wszystkich, którym wyrządził krzywdę.
-
Napisałem do niego kilka dni temu i uważam, że powinniśmy poczekać, aż wejdzie na forum i wyjaśni sprawę, łącznie z przeprosinami. Jeśli nie, to zawsze jest czas na nałożenie bana permanentnego.
-
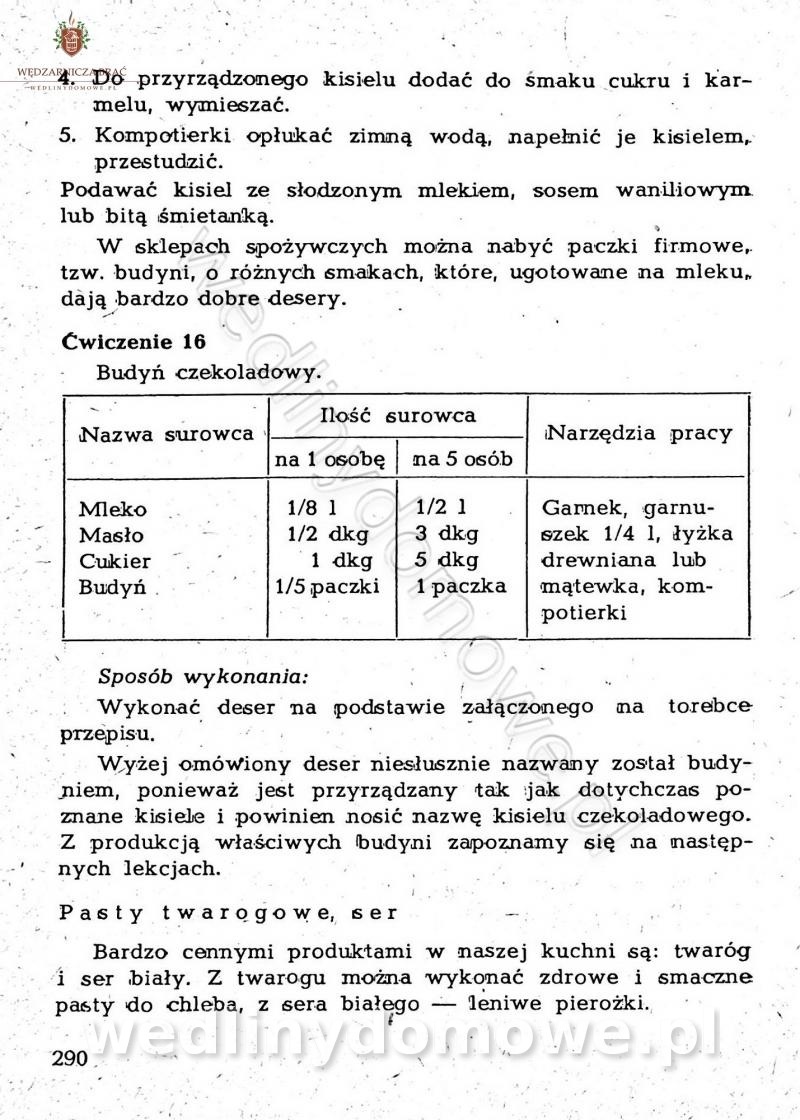
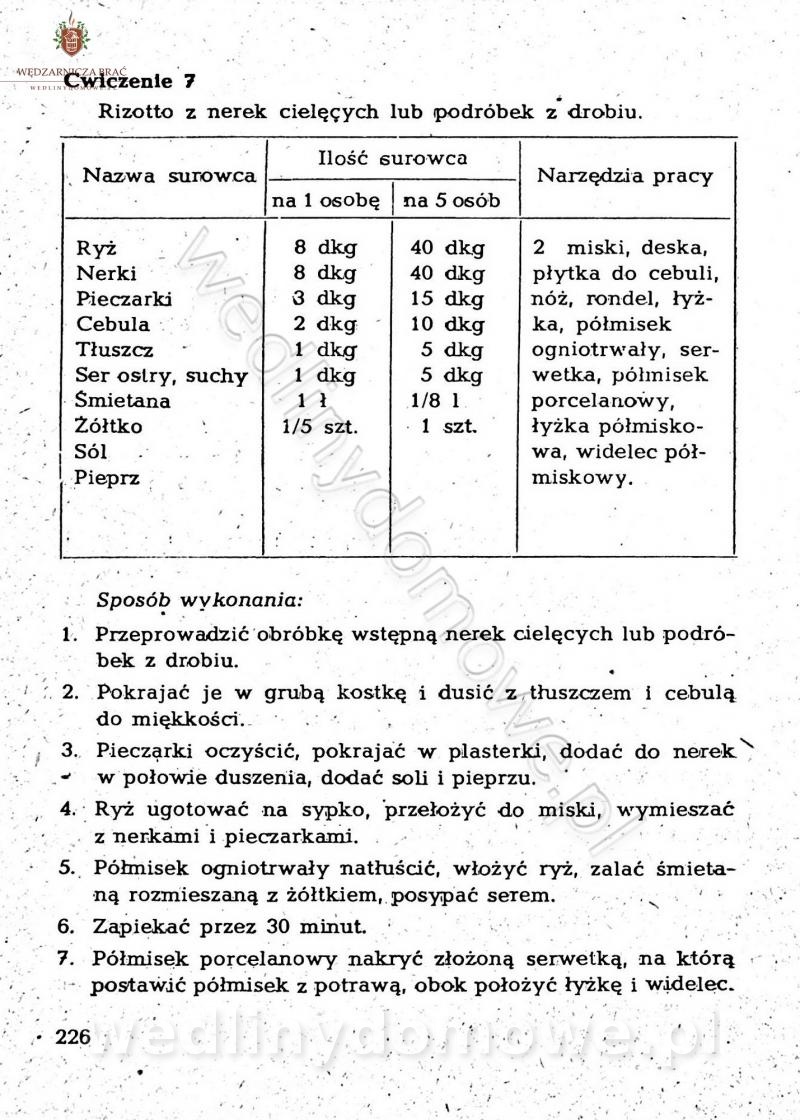
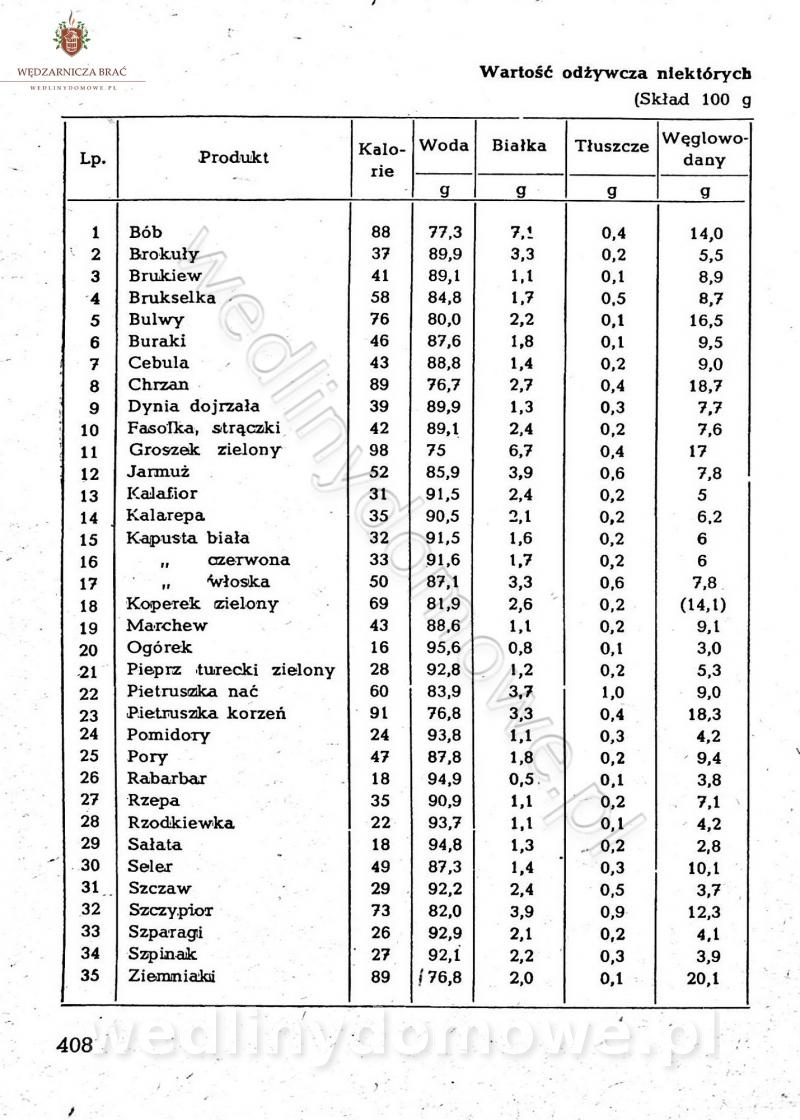
[Rok 1958][Tom 2] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie

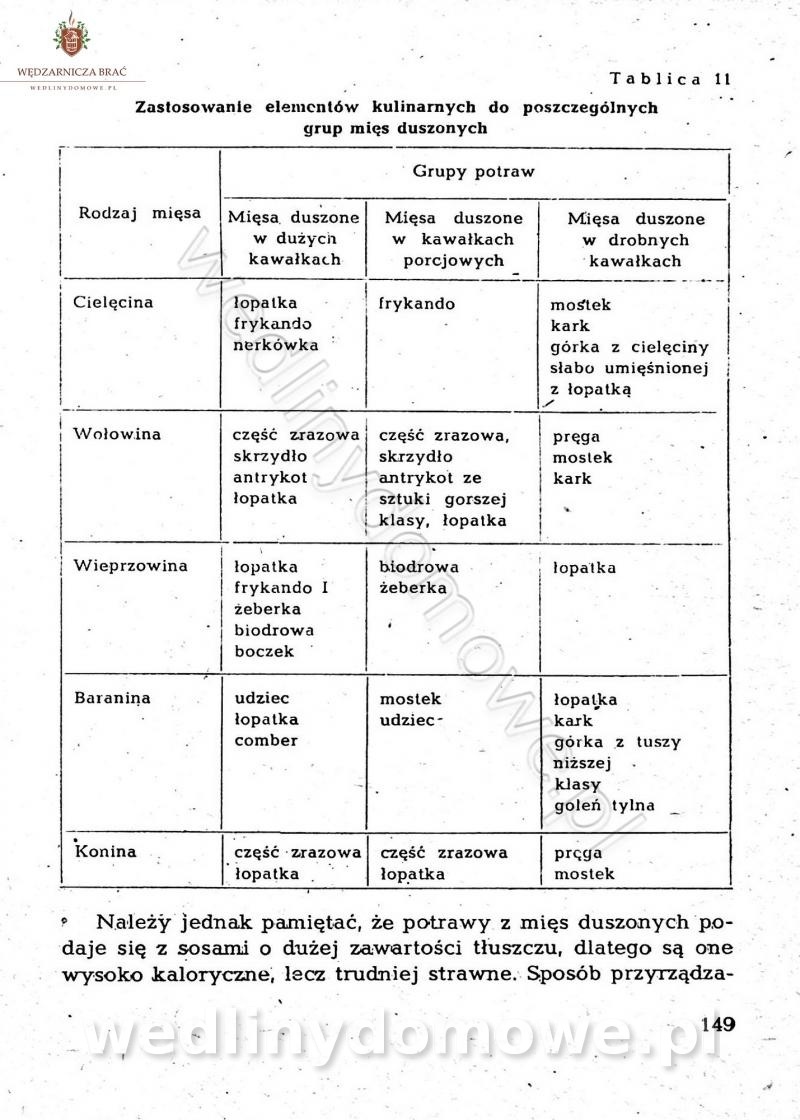



























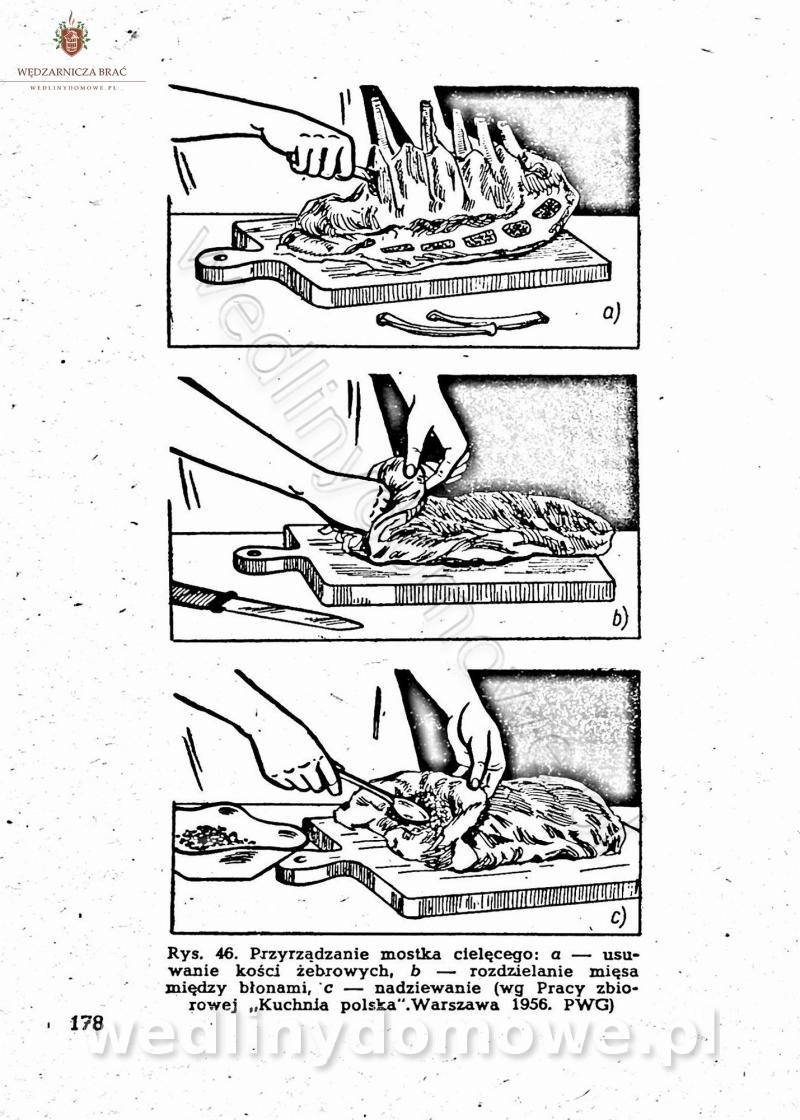
















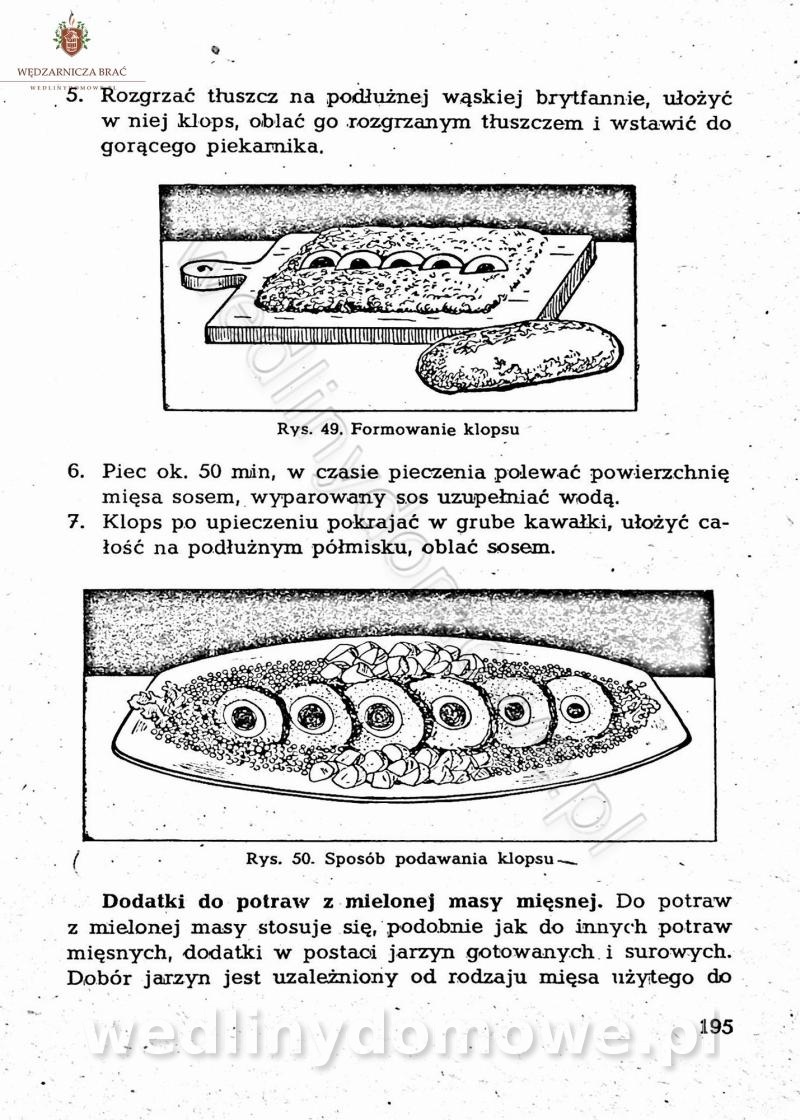
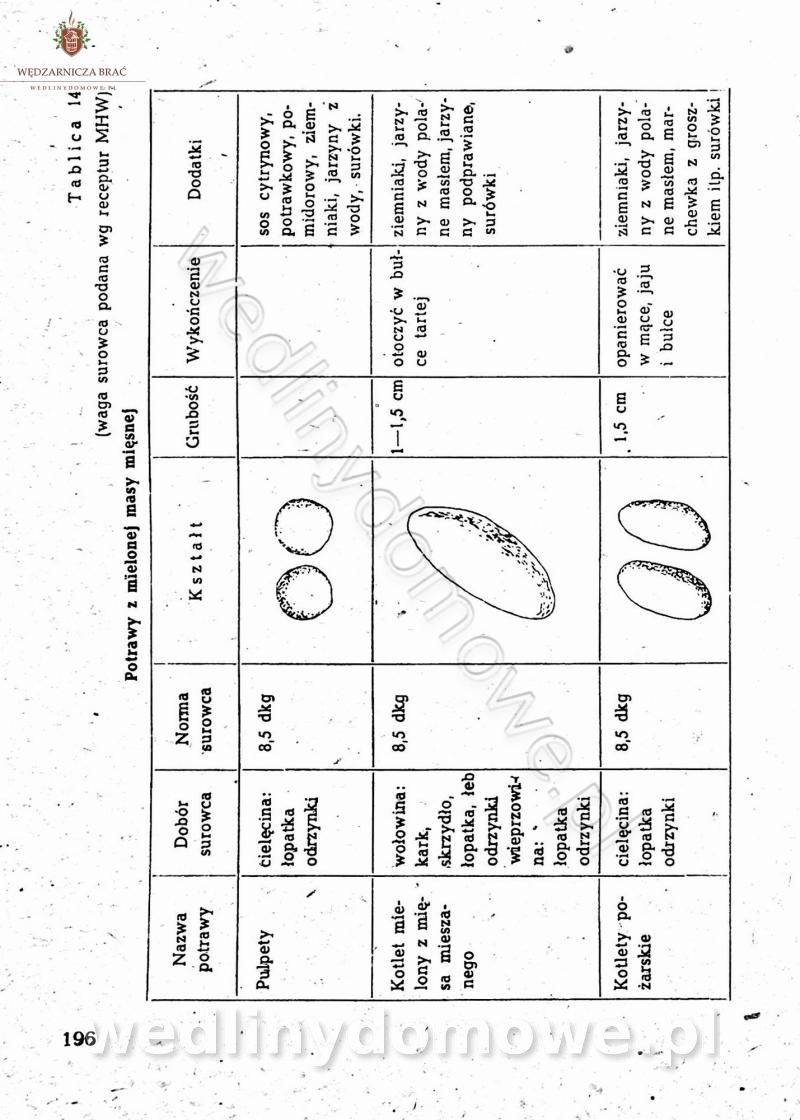
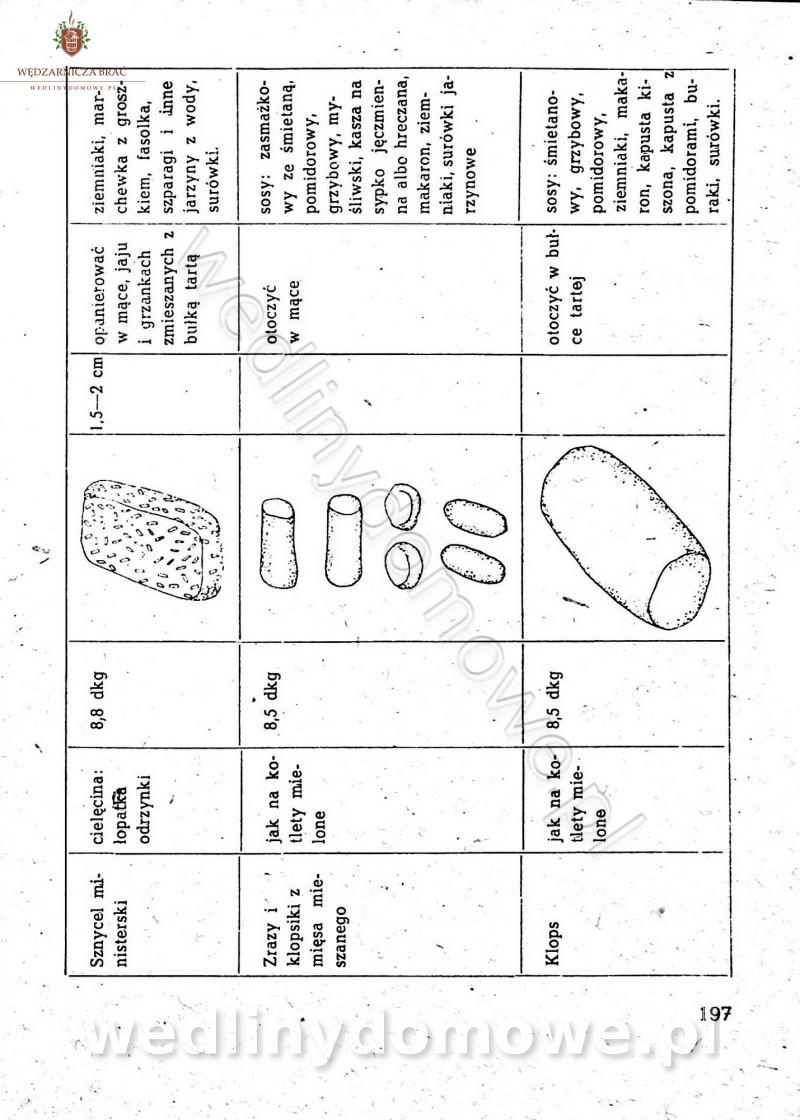
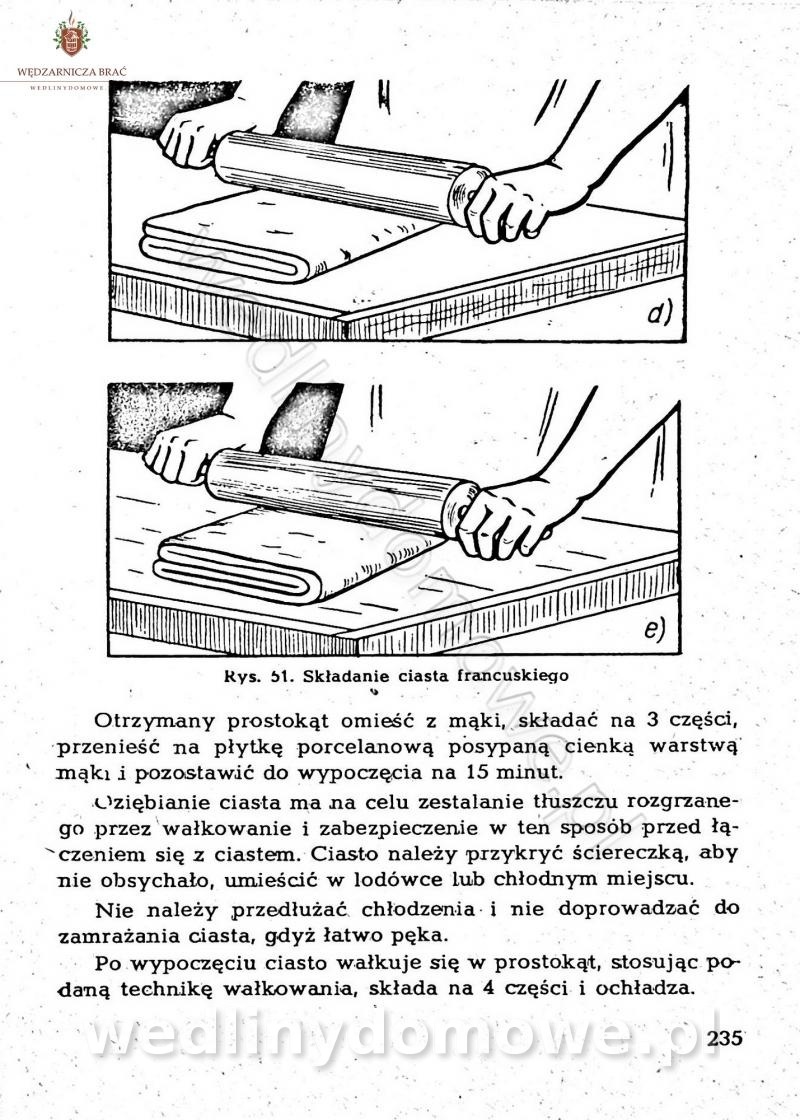
-
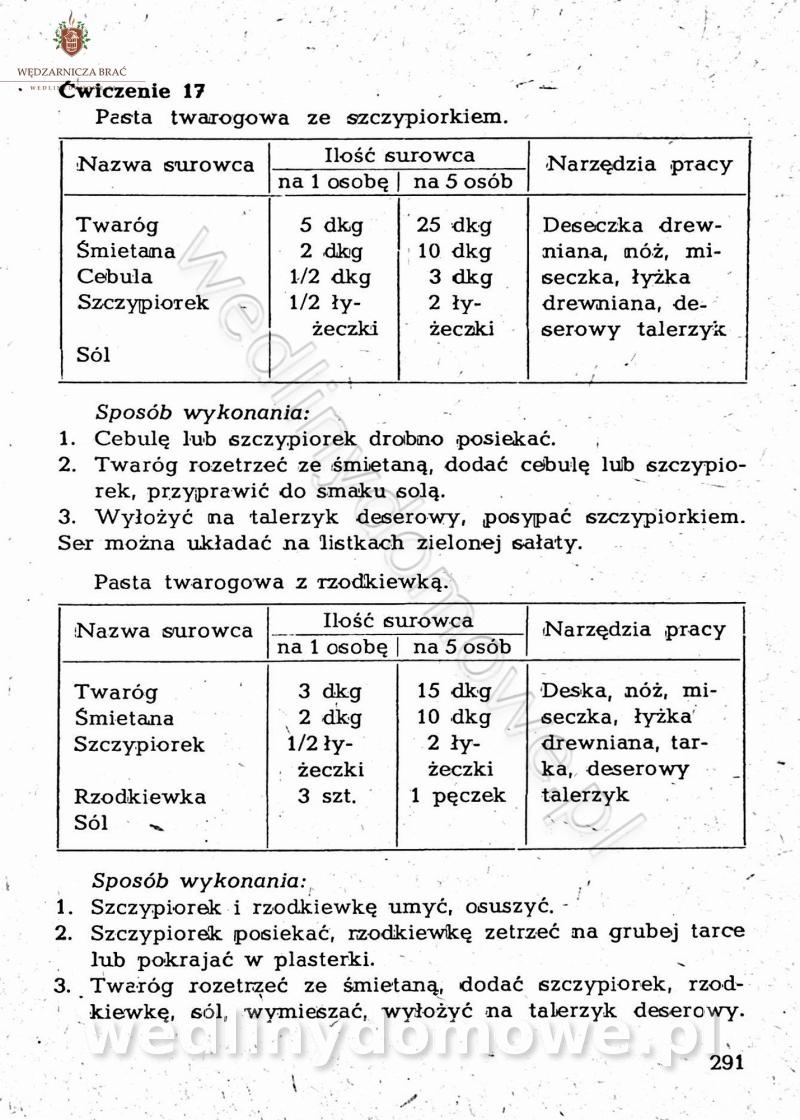
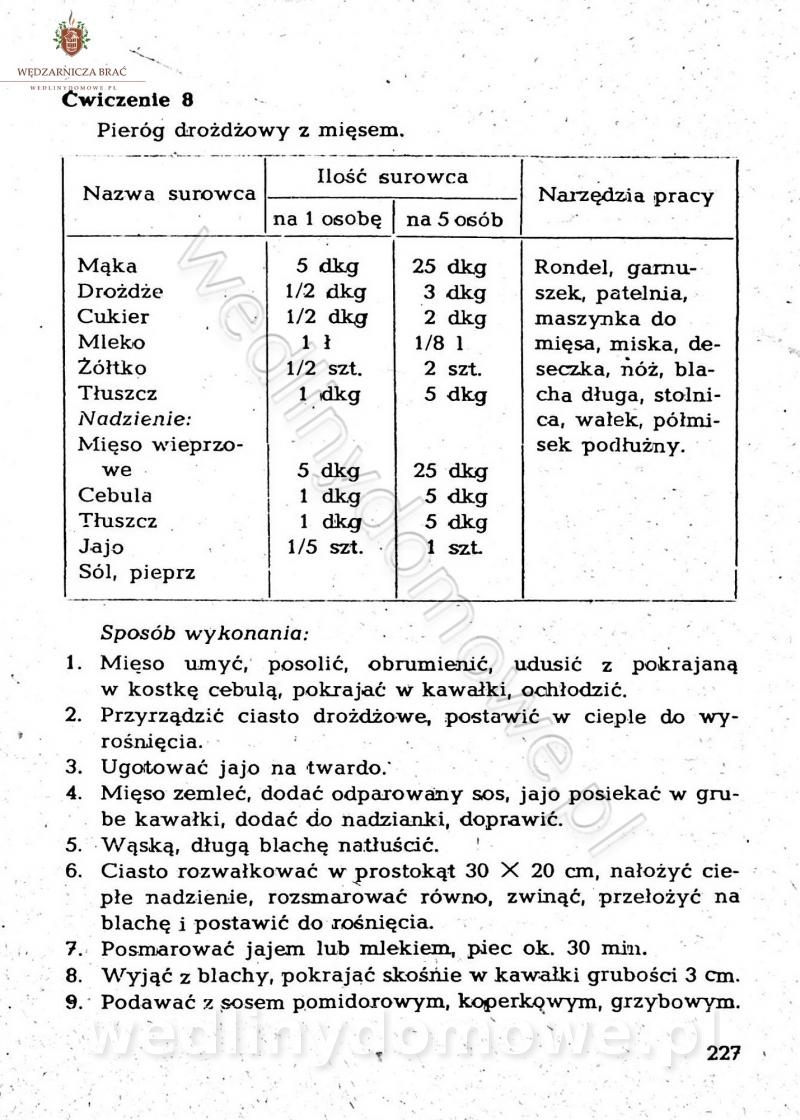
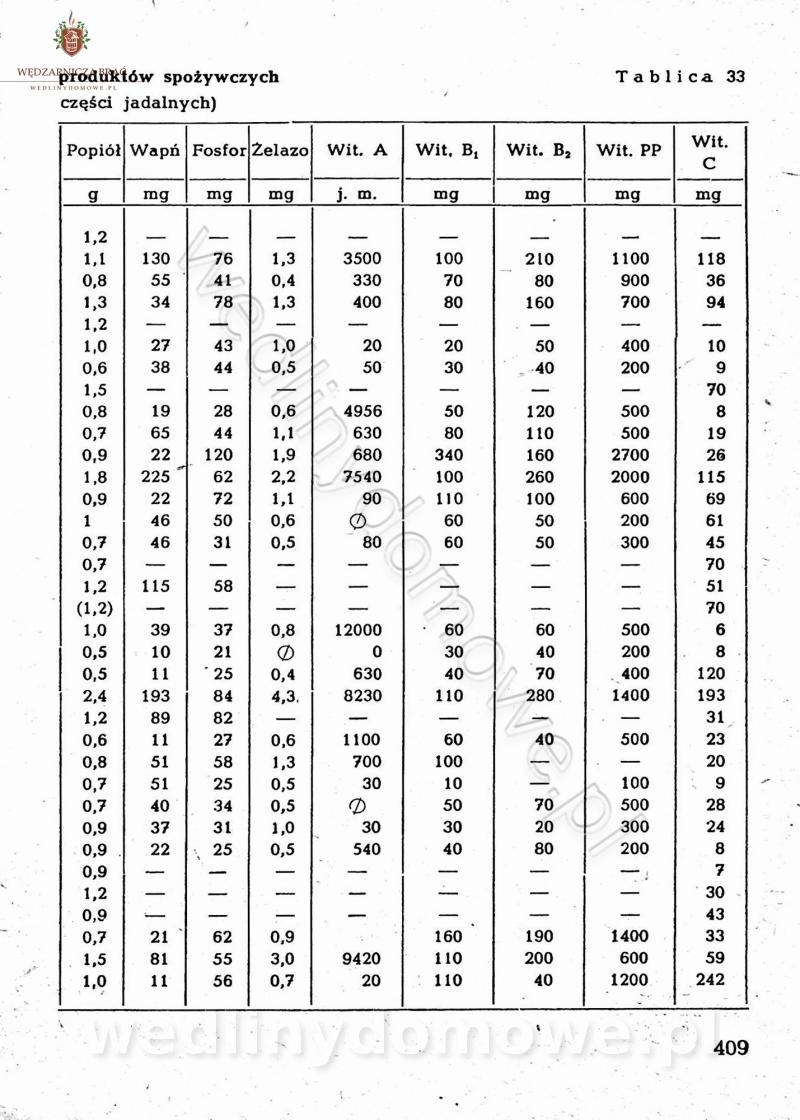
[Rok 1958][Tom 2] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie



























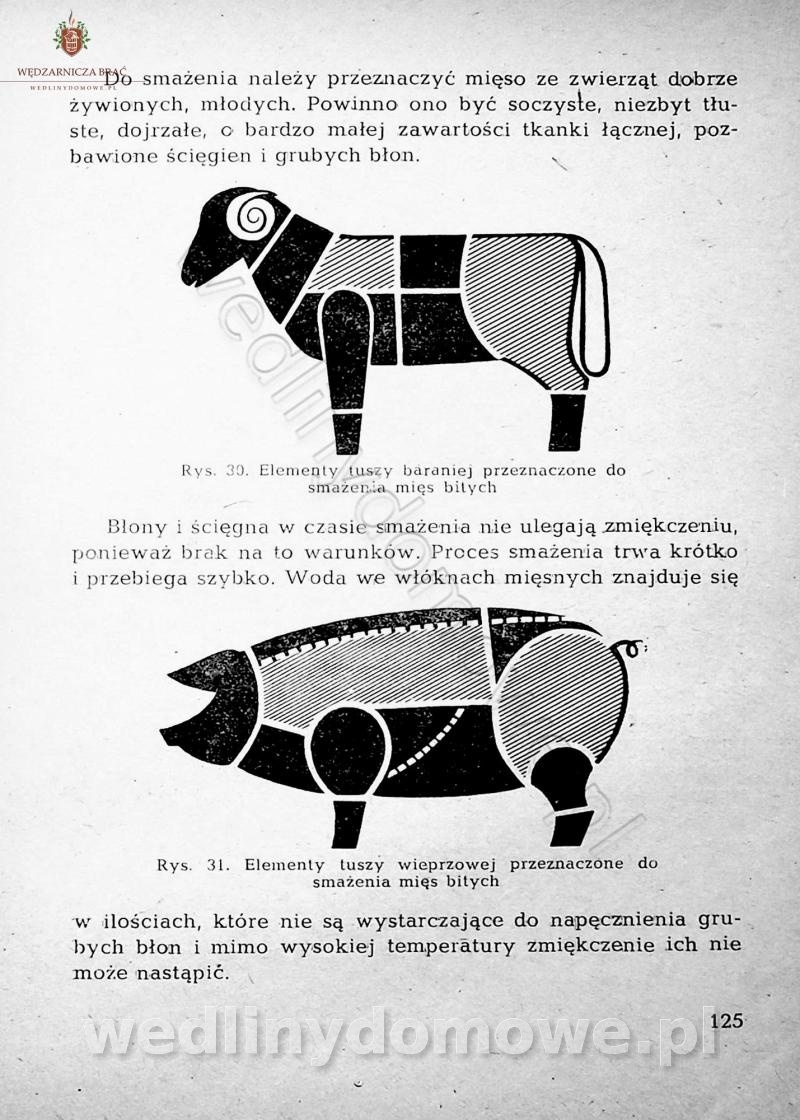

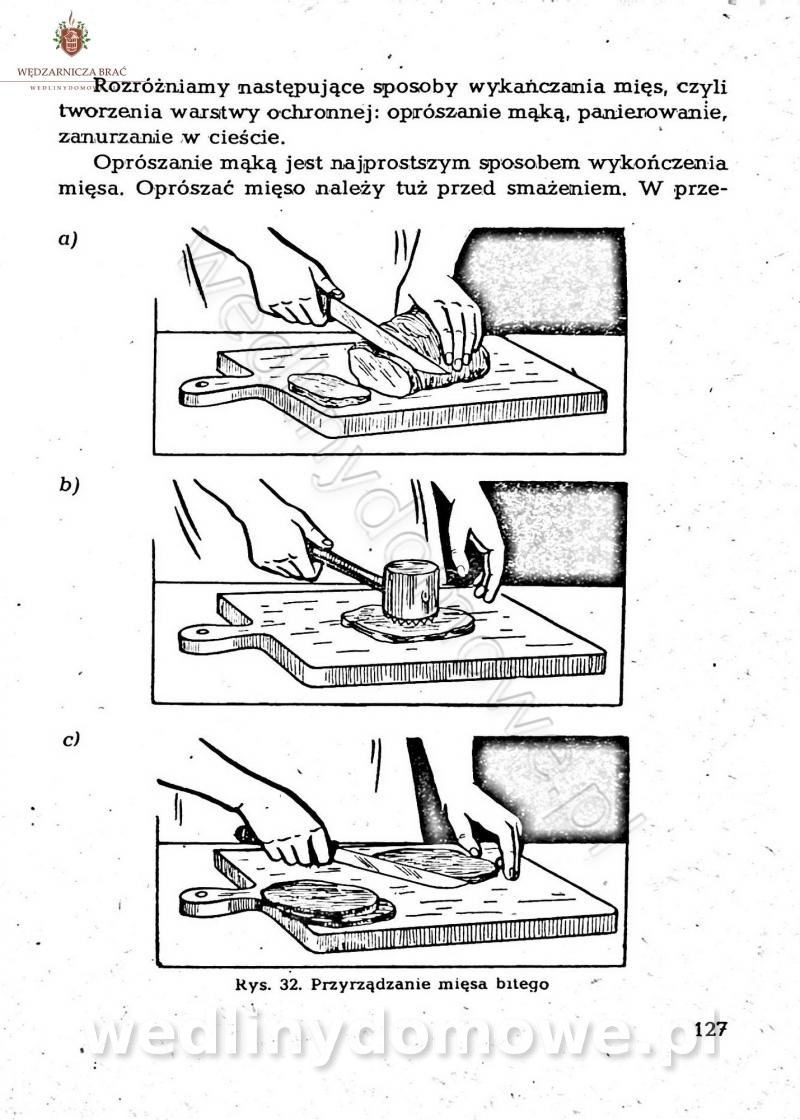


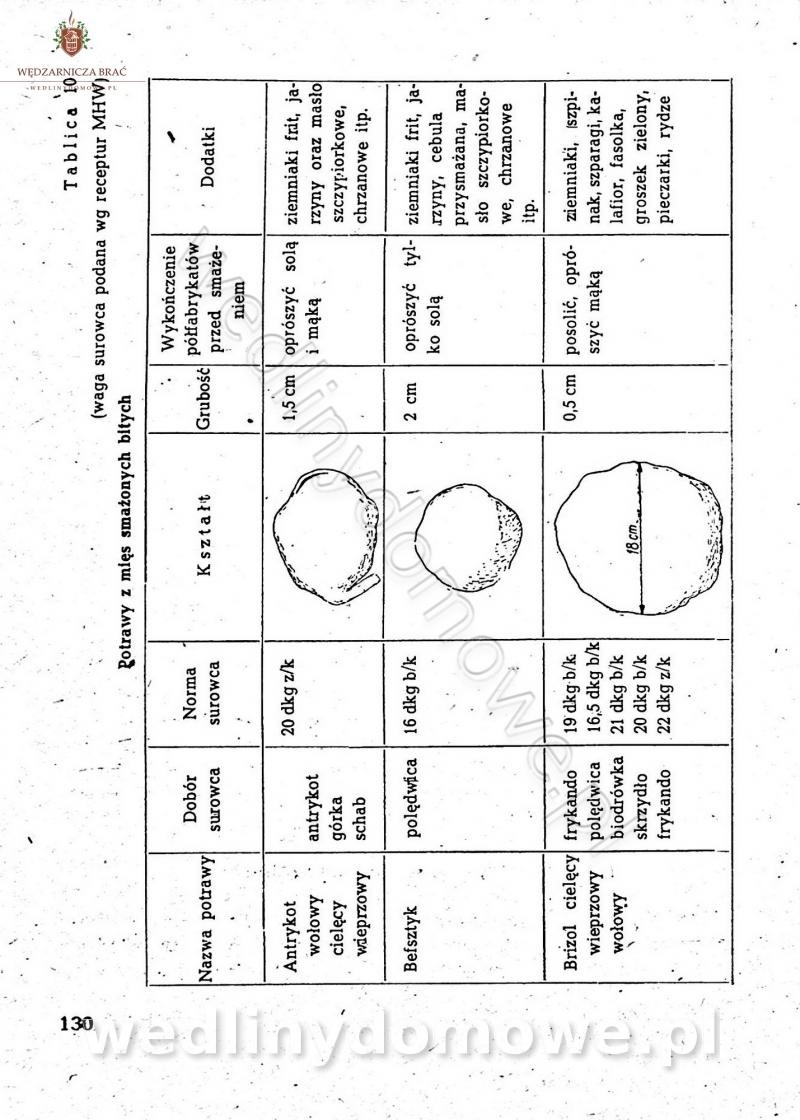
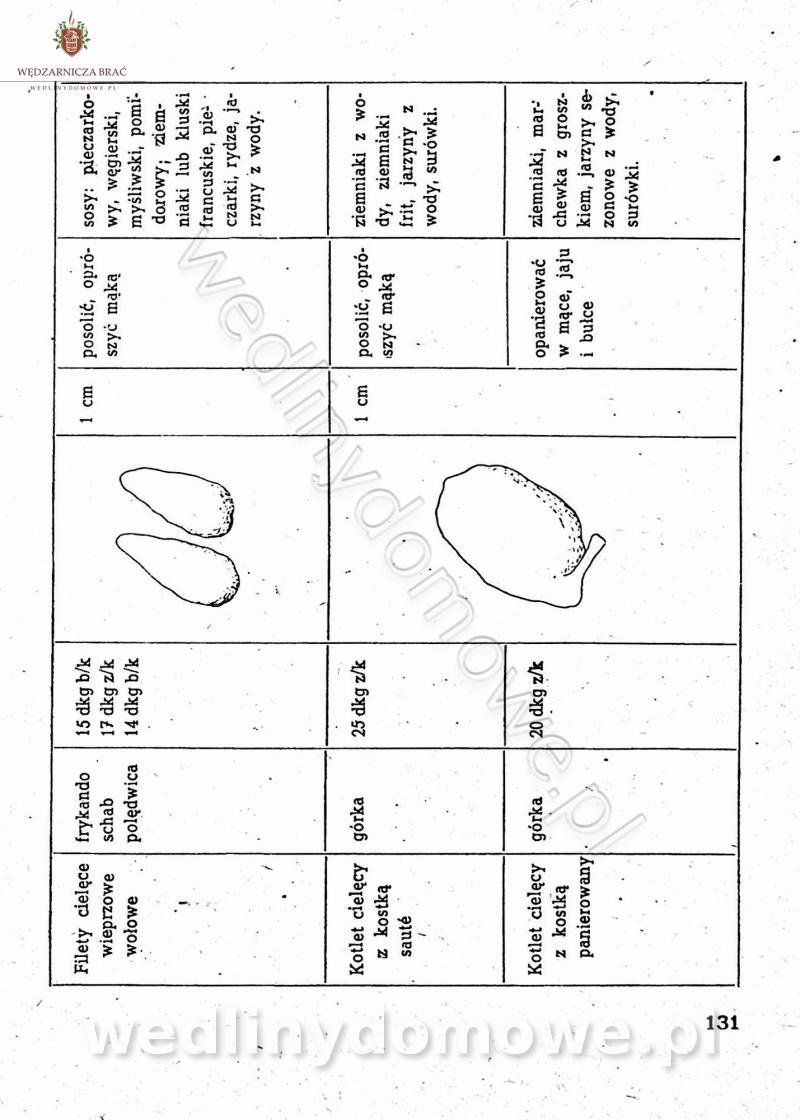
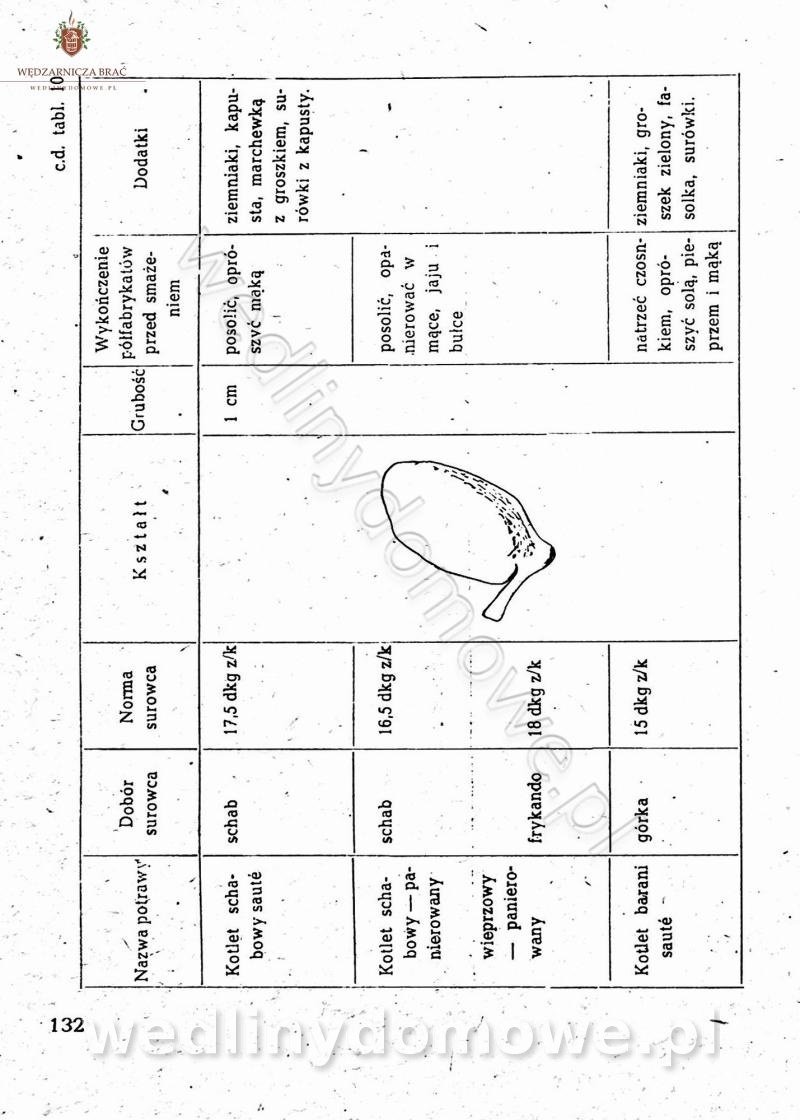
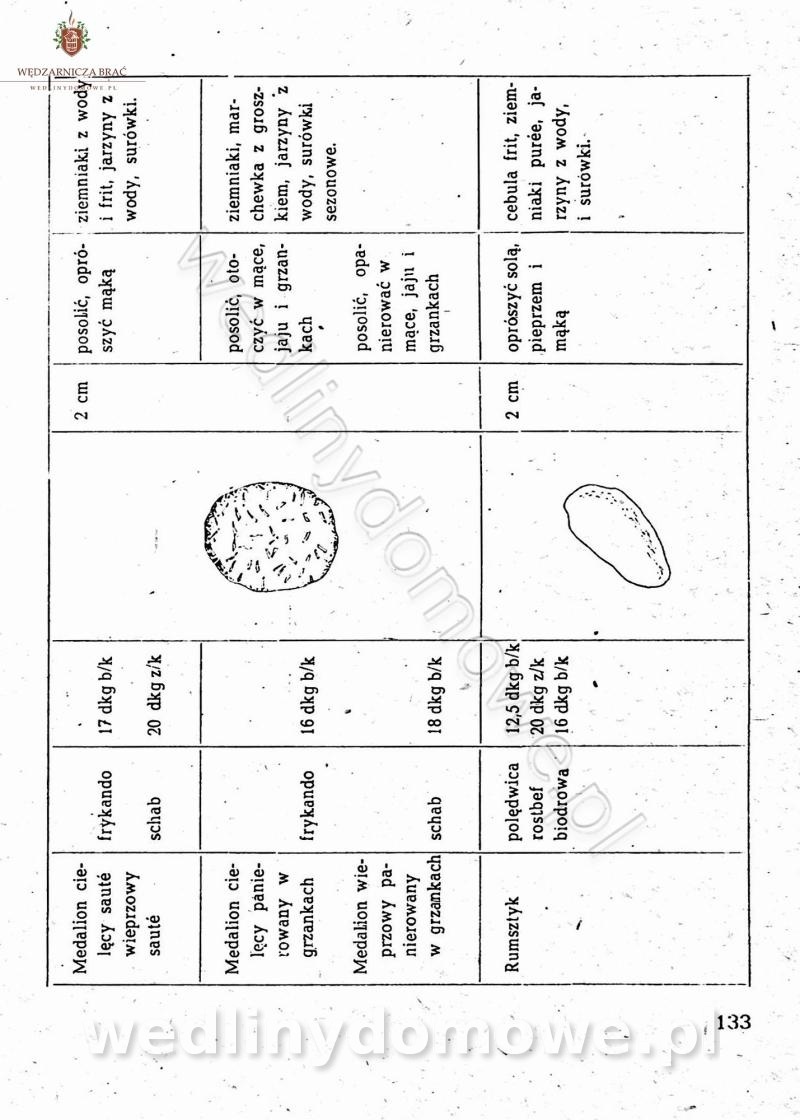
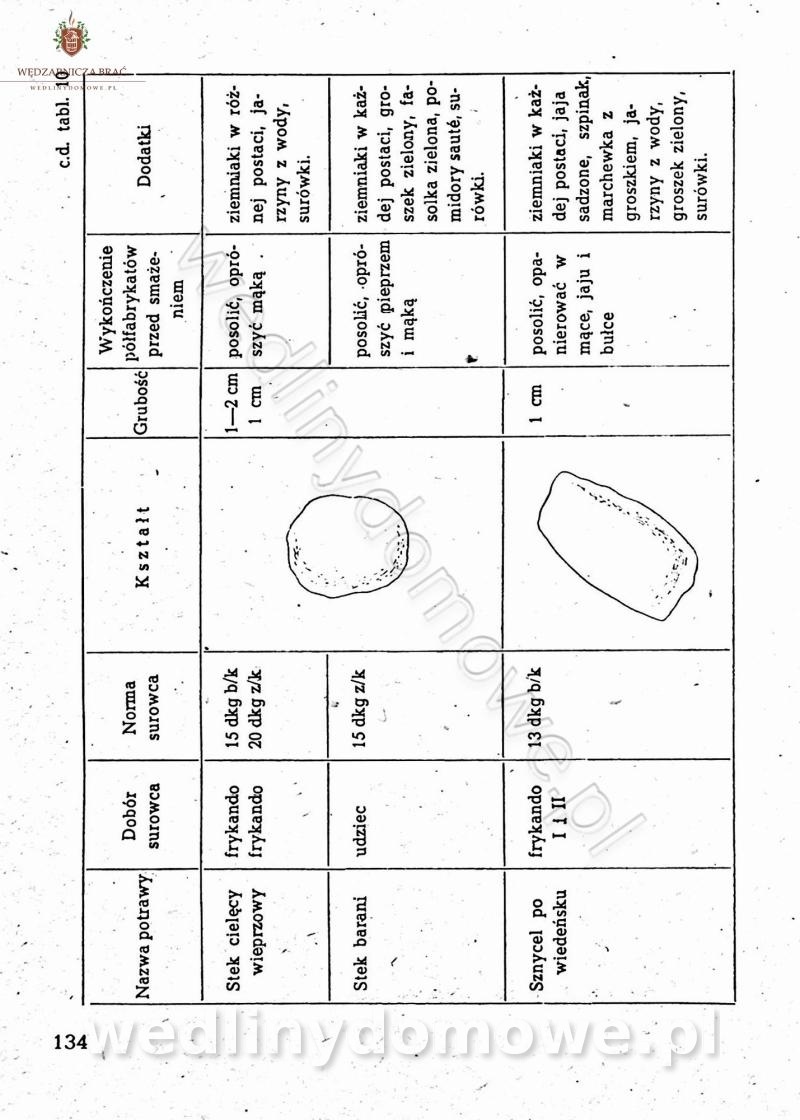








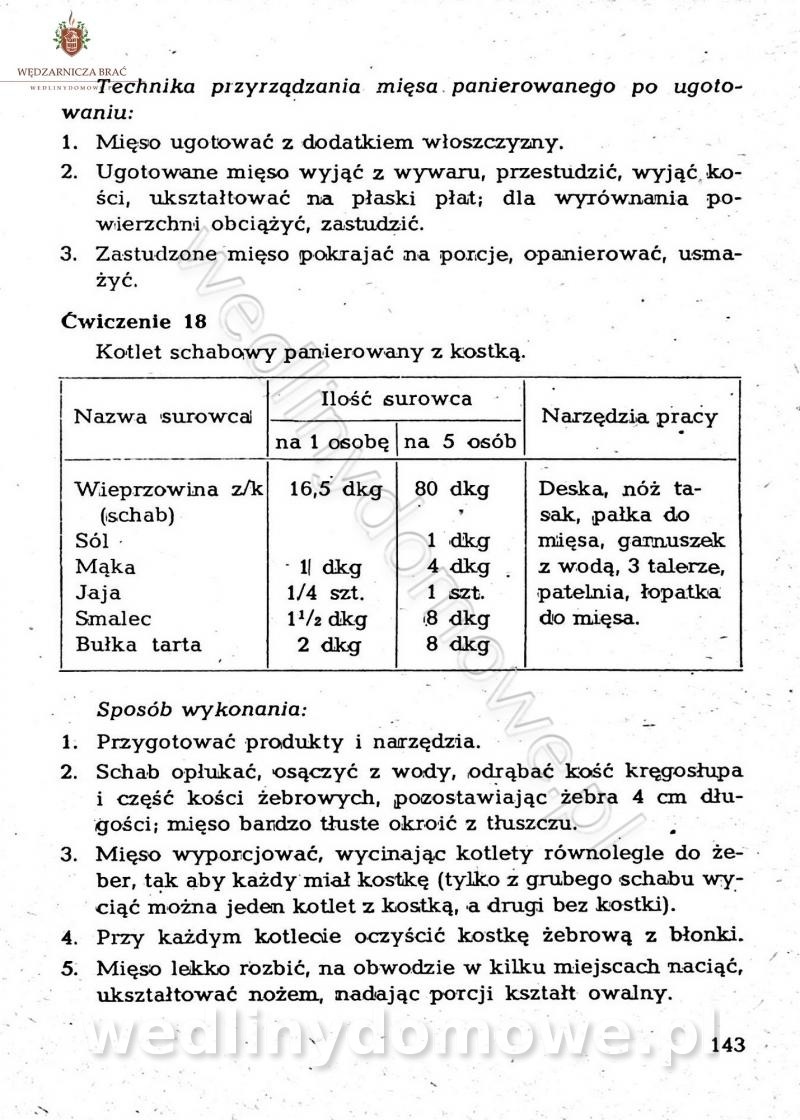

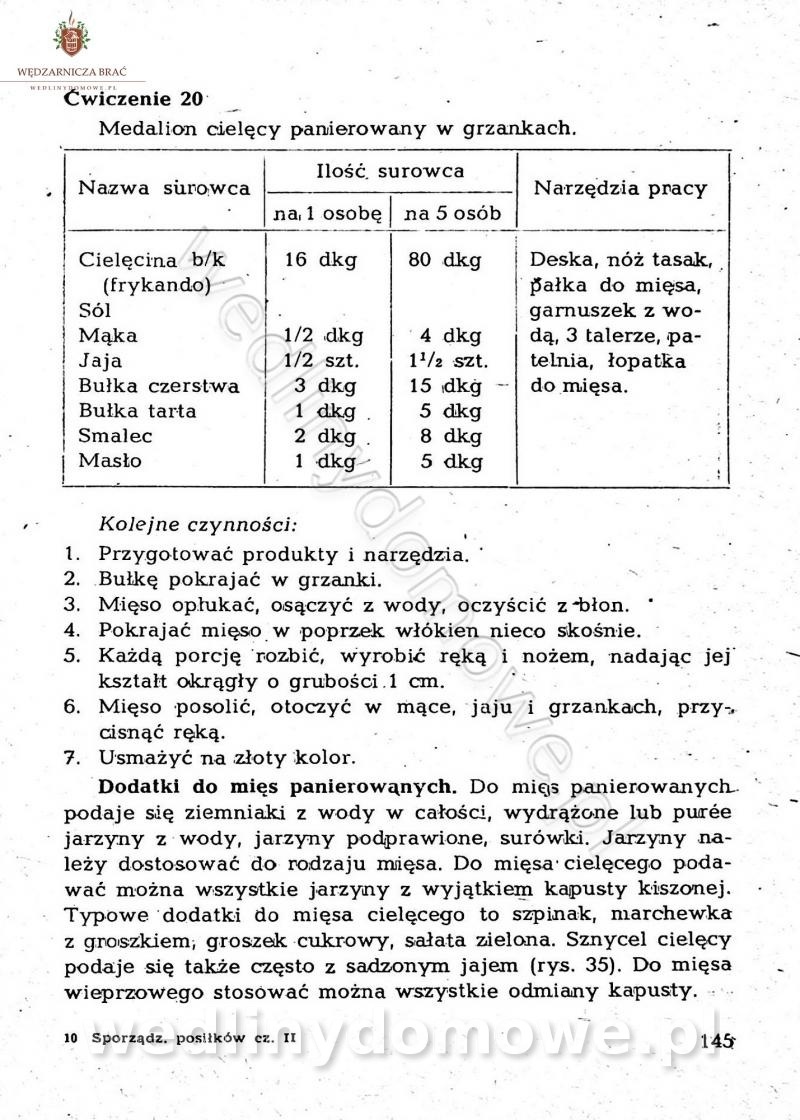
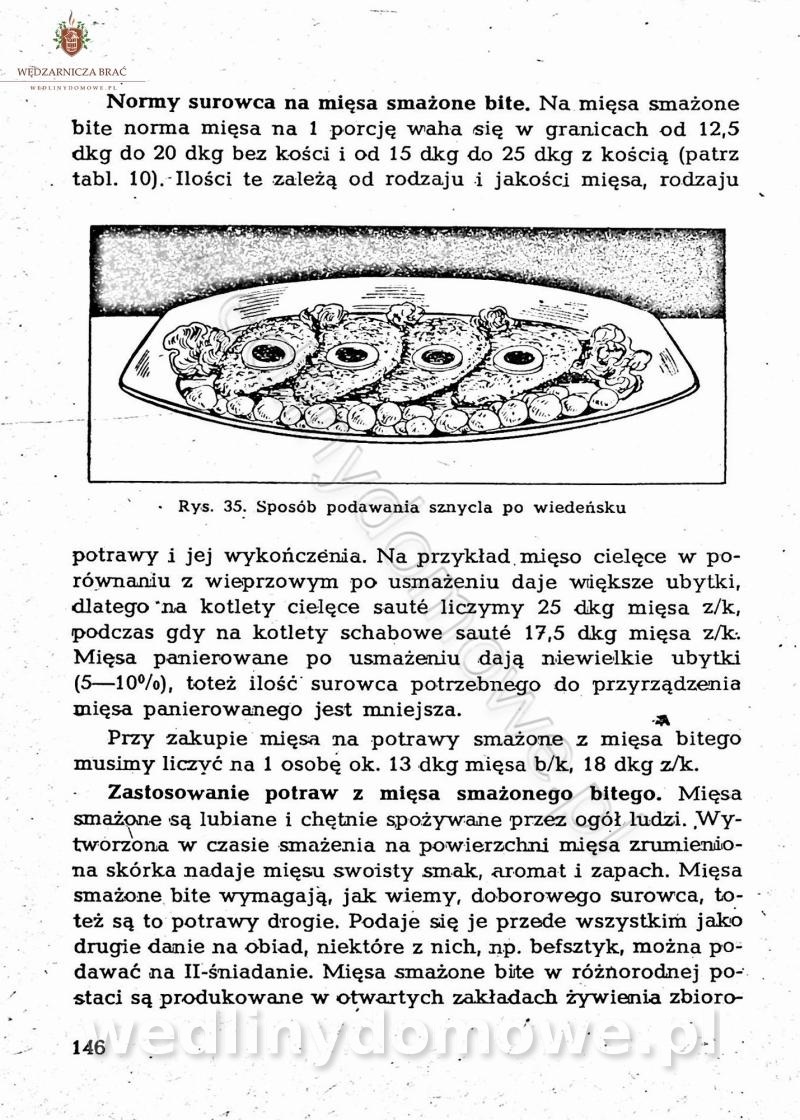

-
Napisałem do @MlKl, aby wypowiedział się w temacie i zamknął go wreszcie.
-
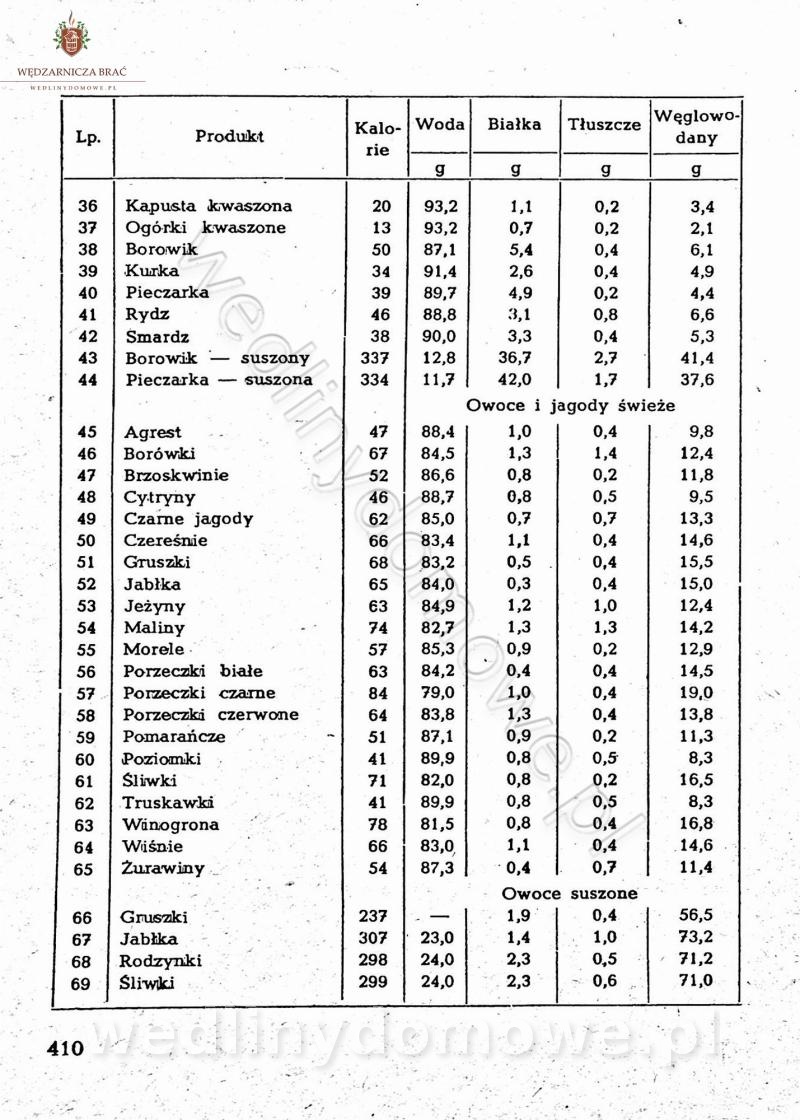
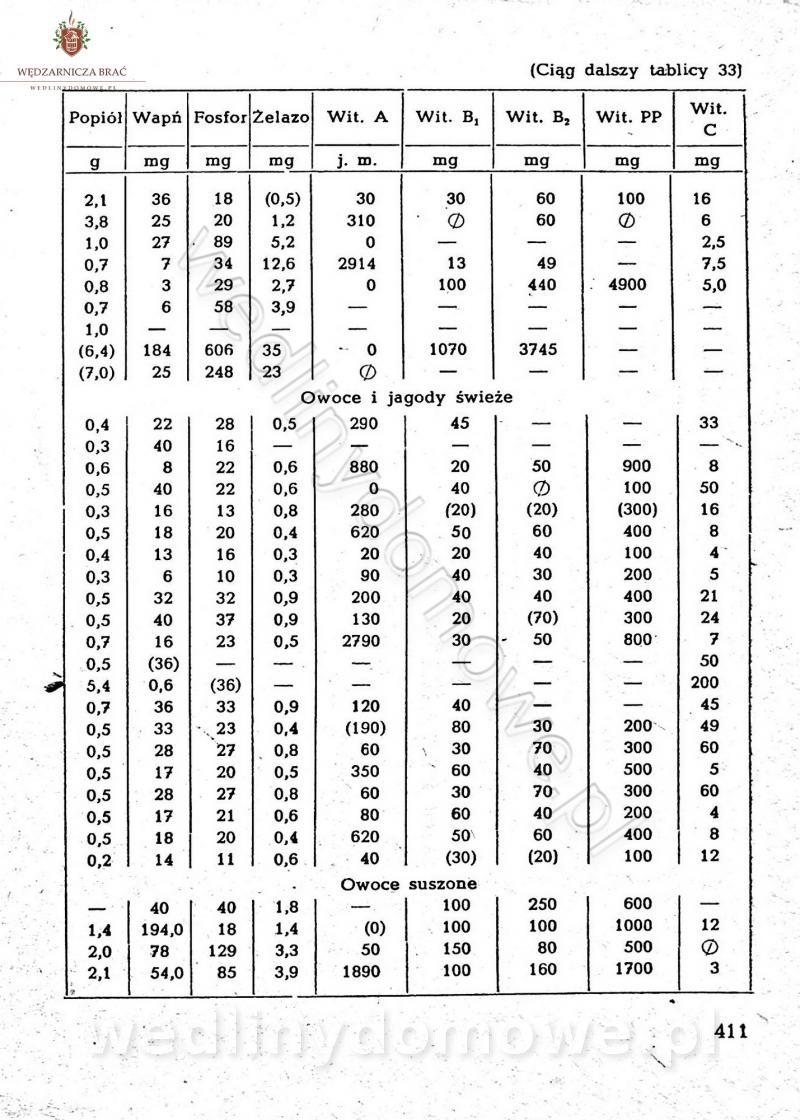
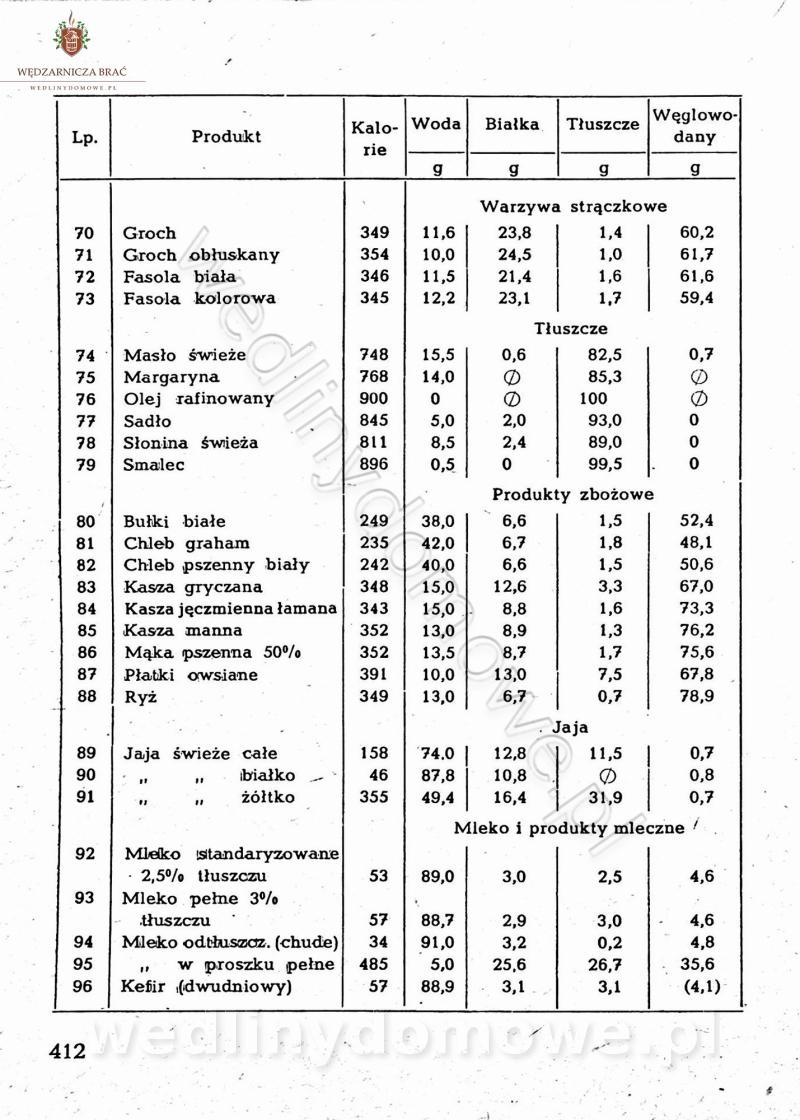
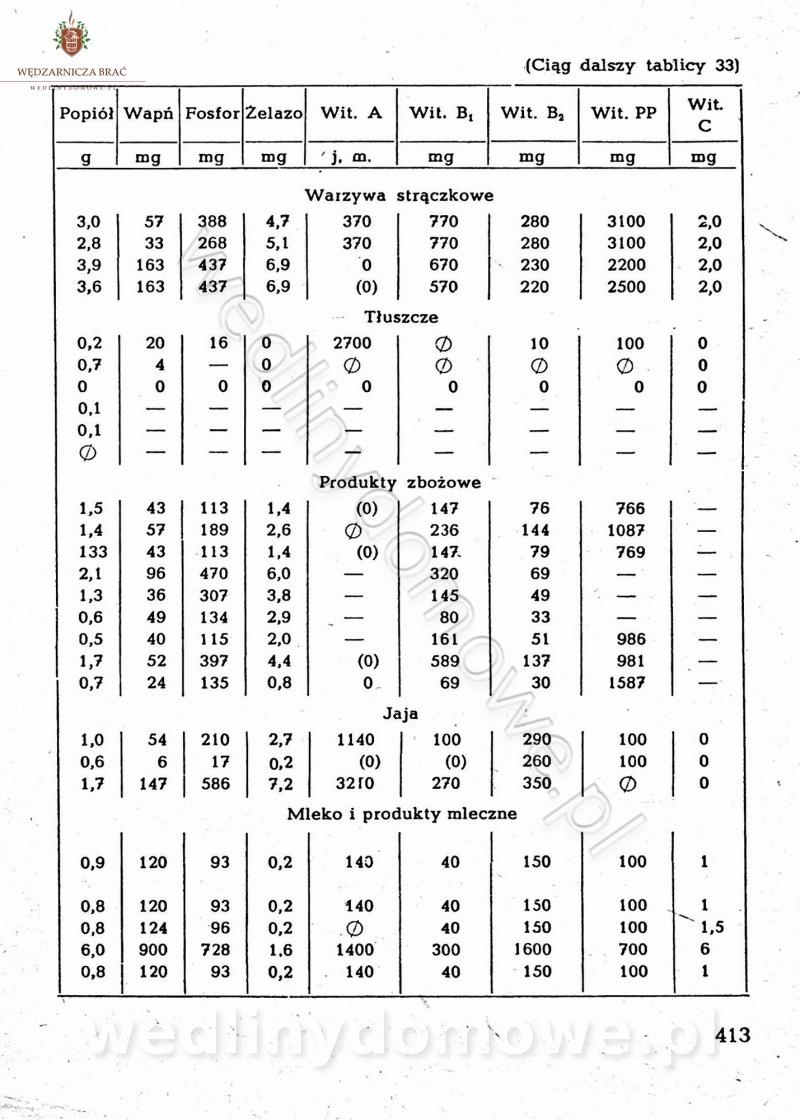
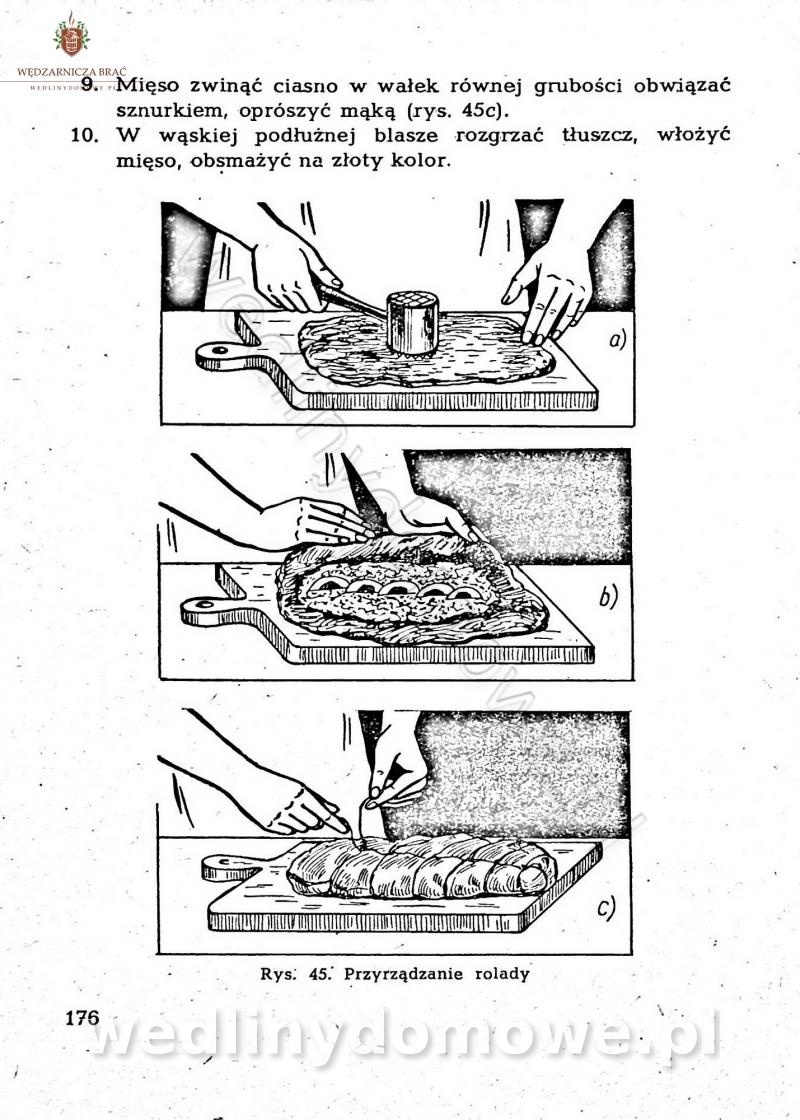
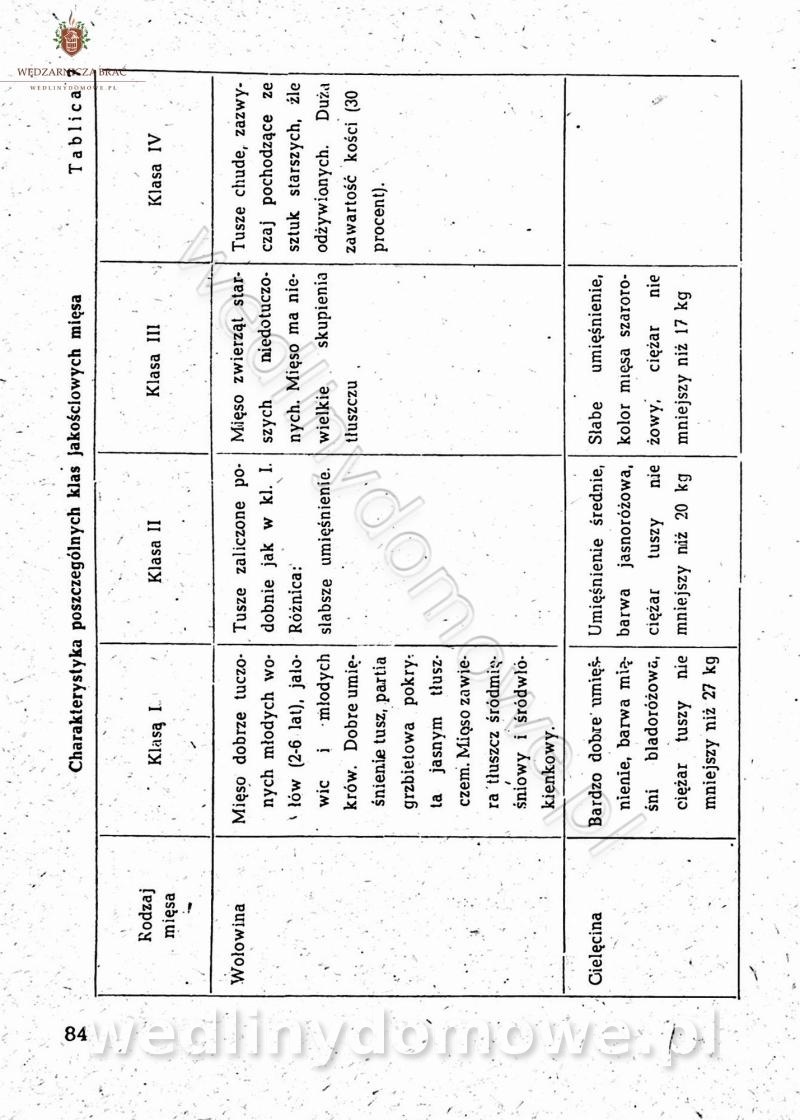
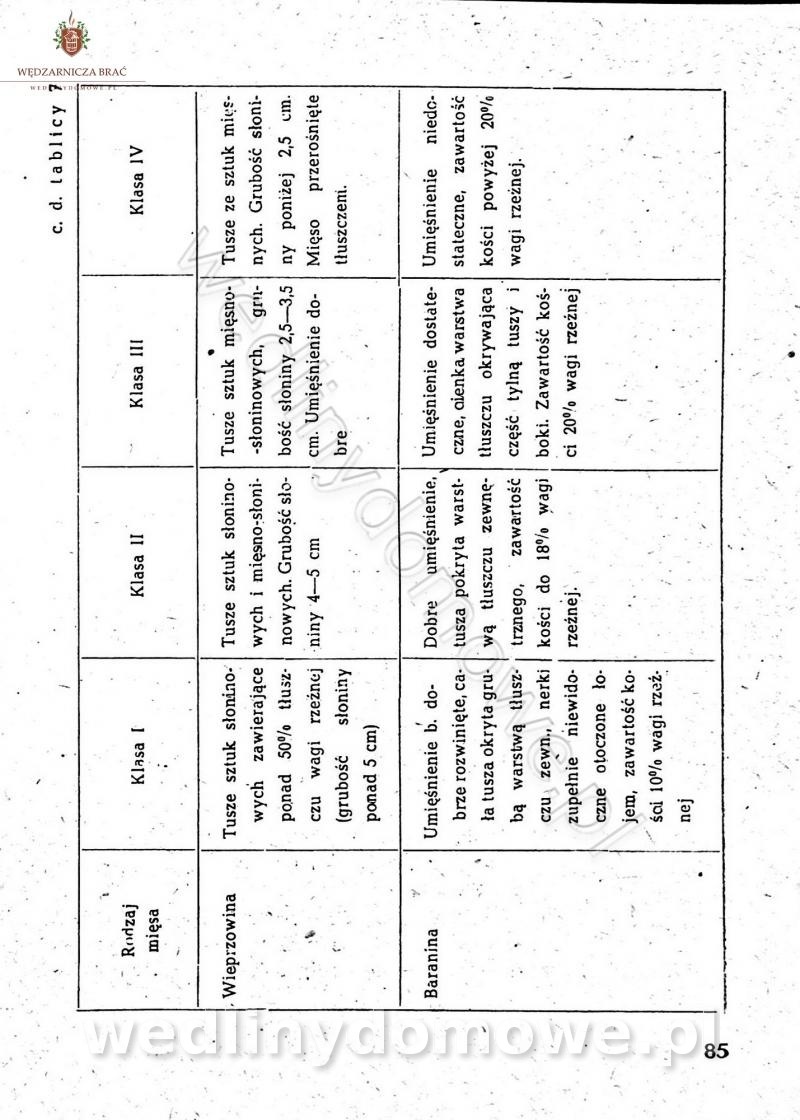
[Rok 1958][Tom 2] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie



















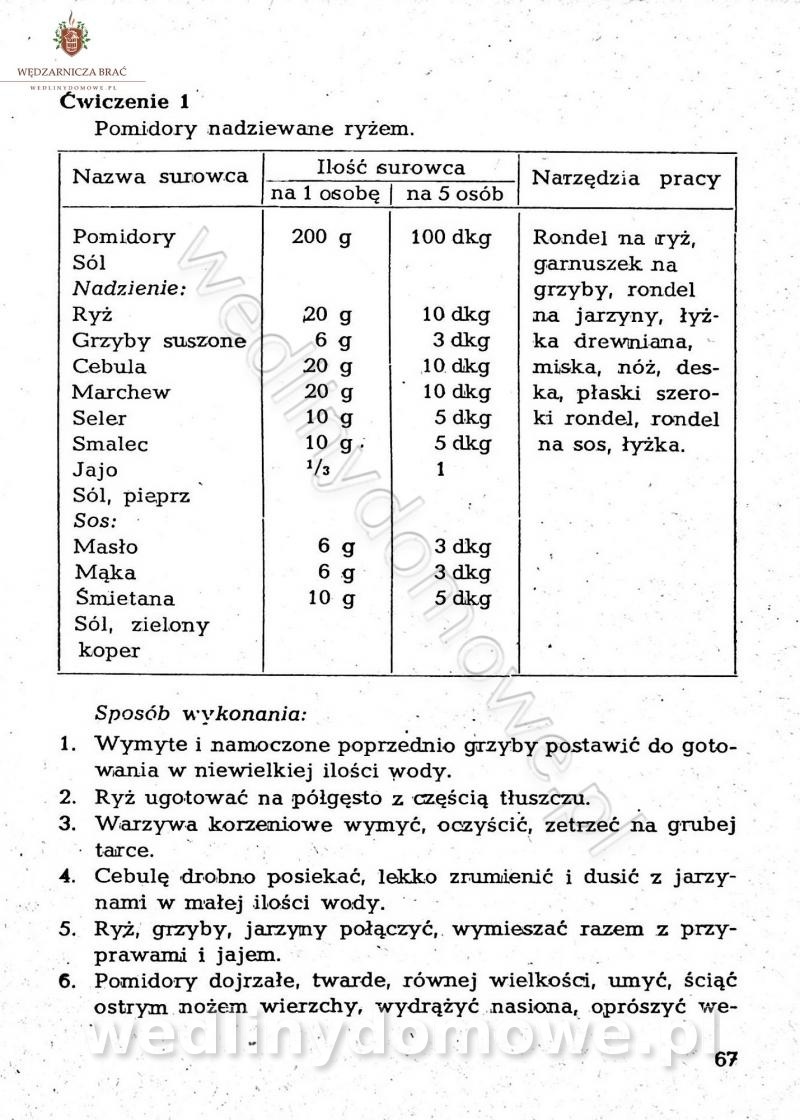


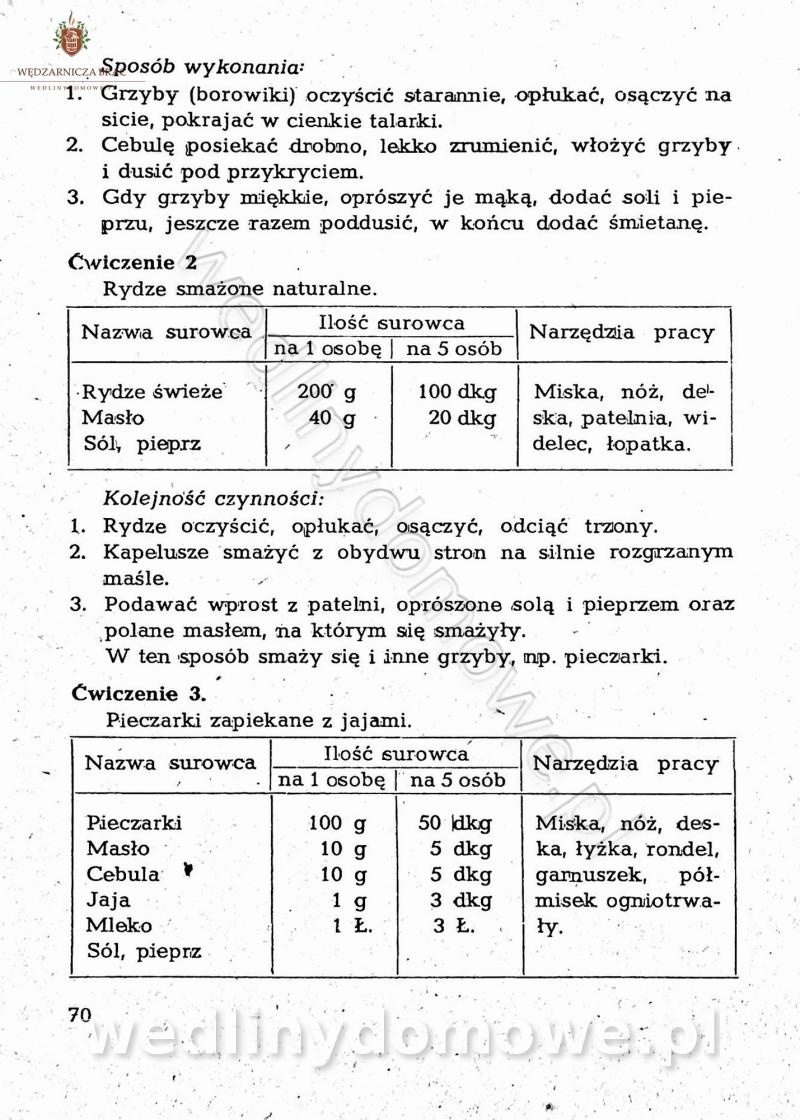
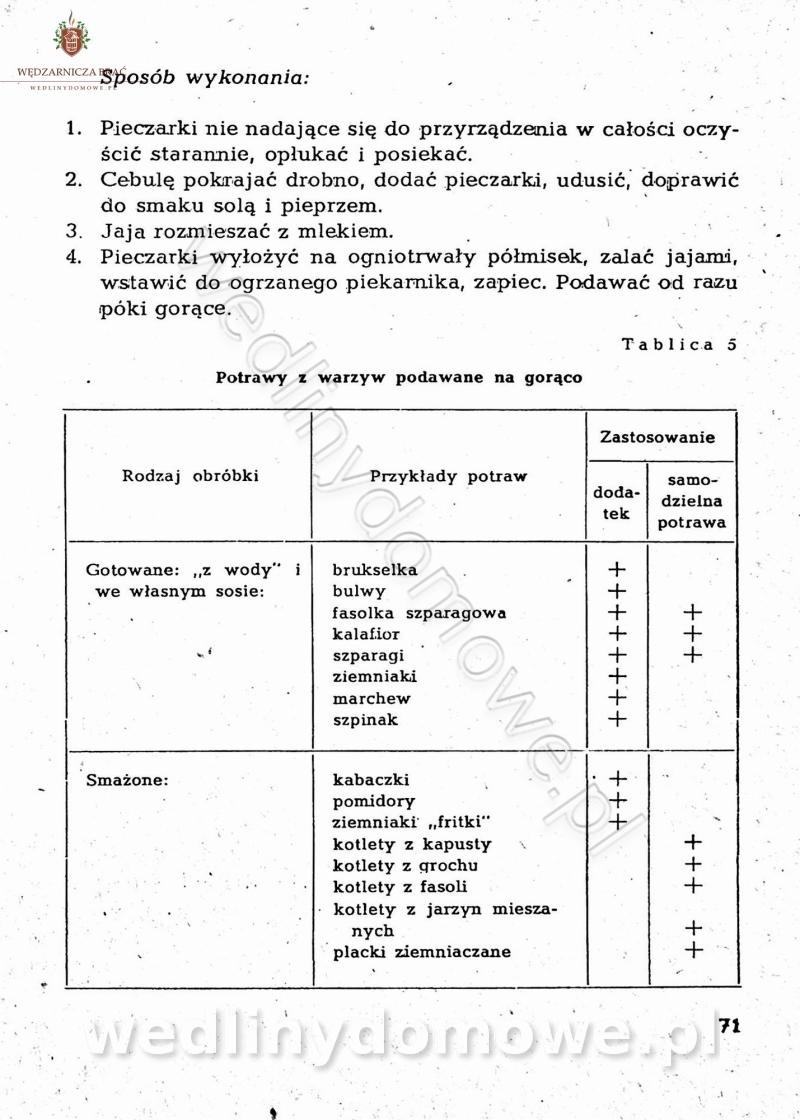
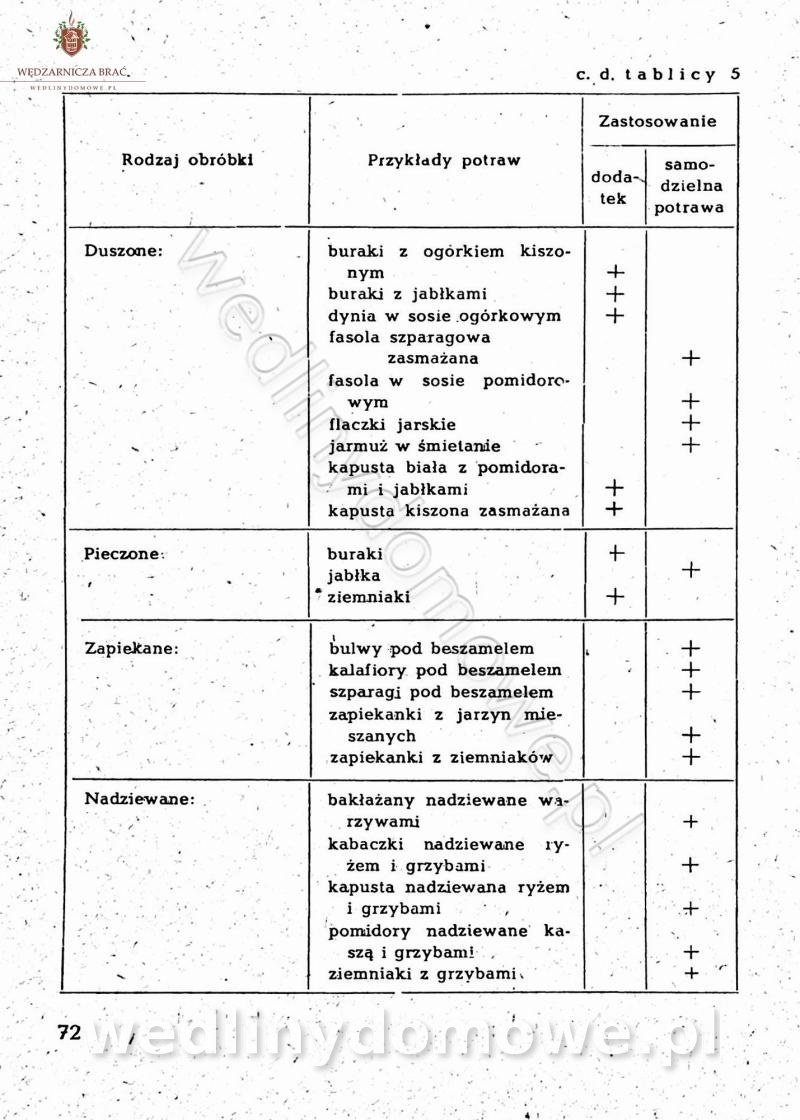


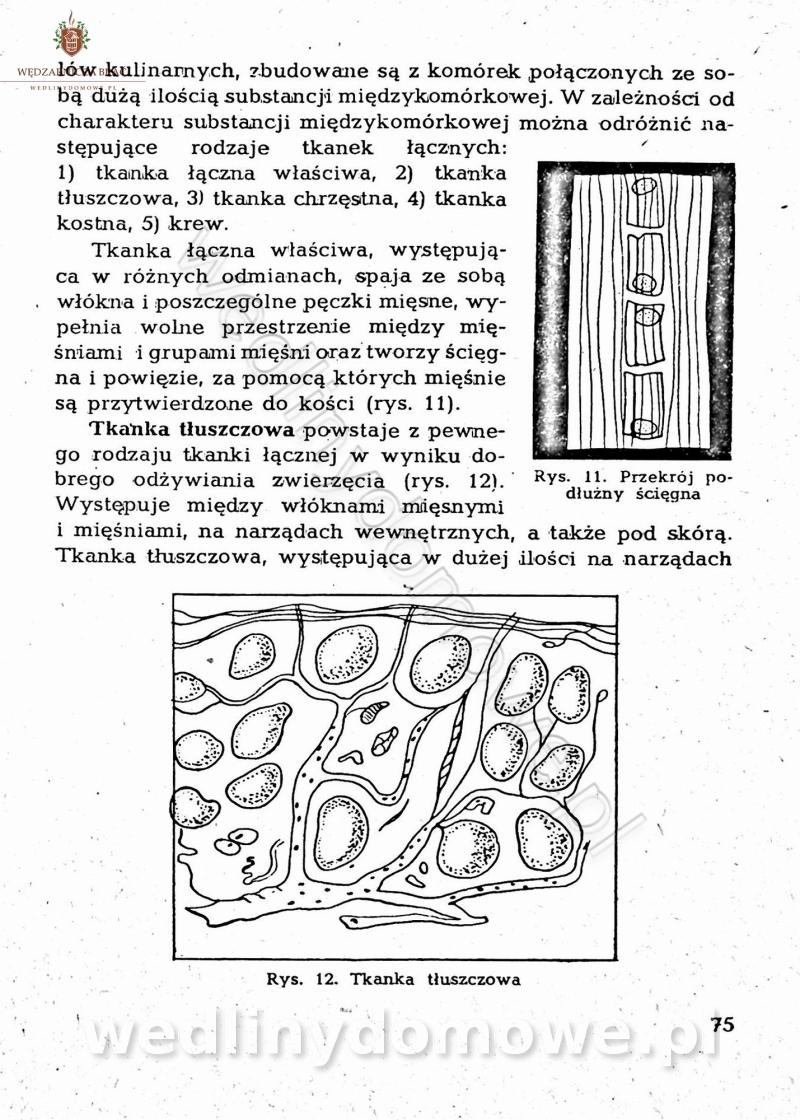




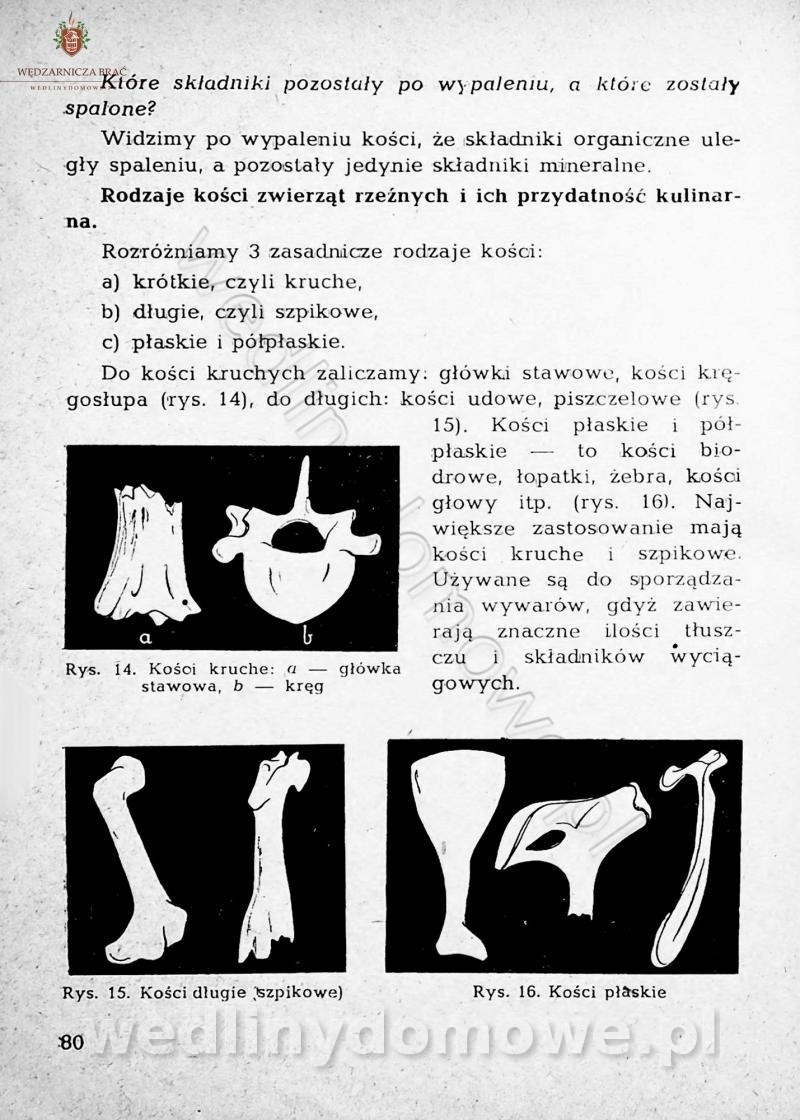















-























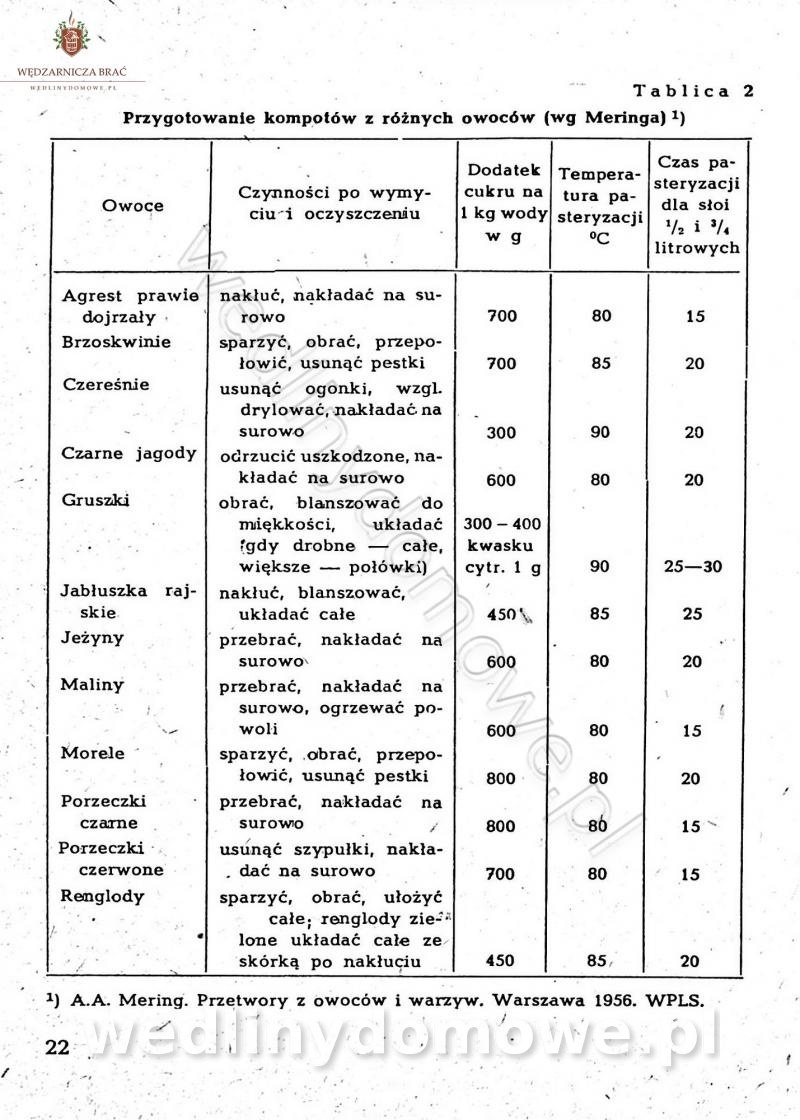
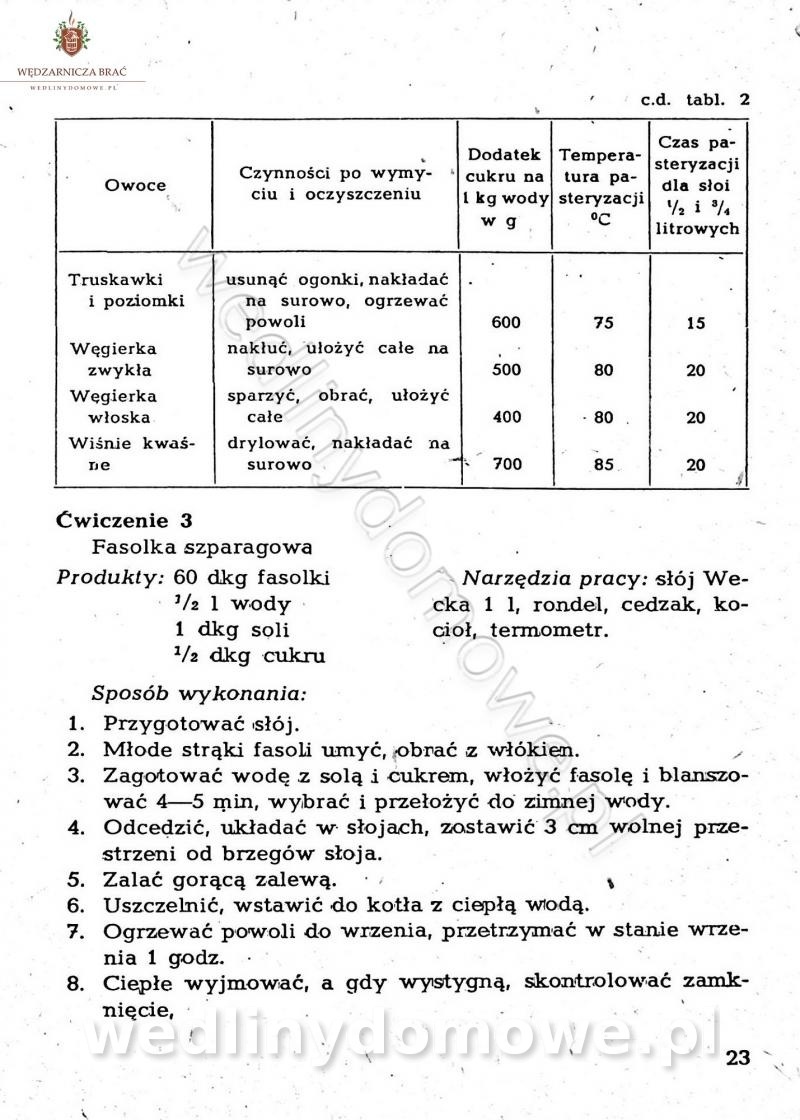









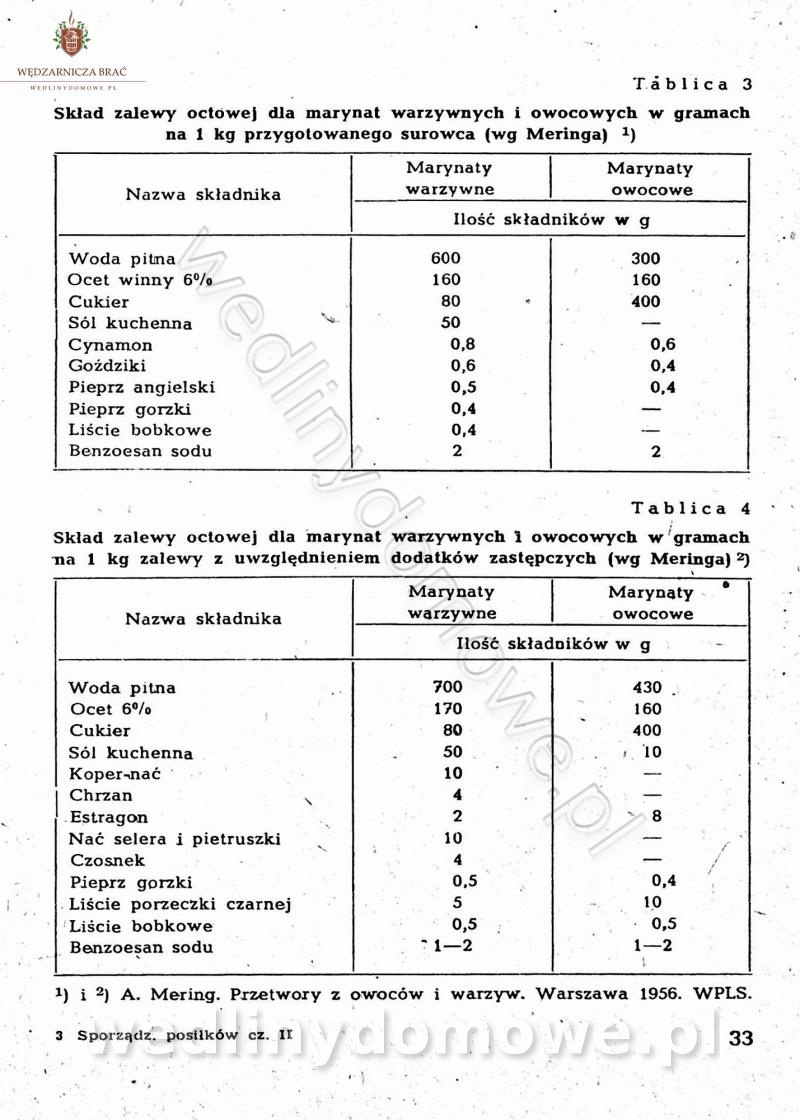














-
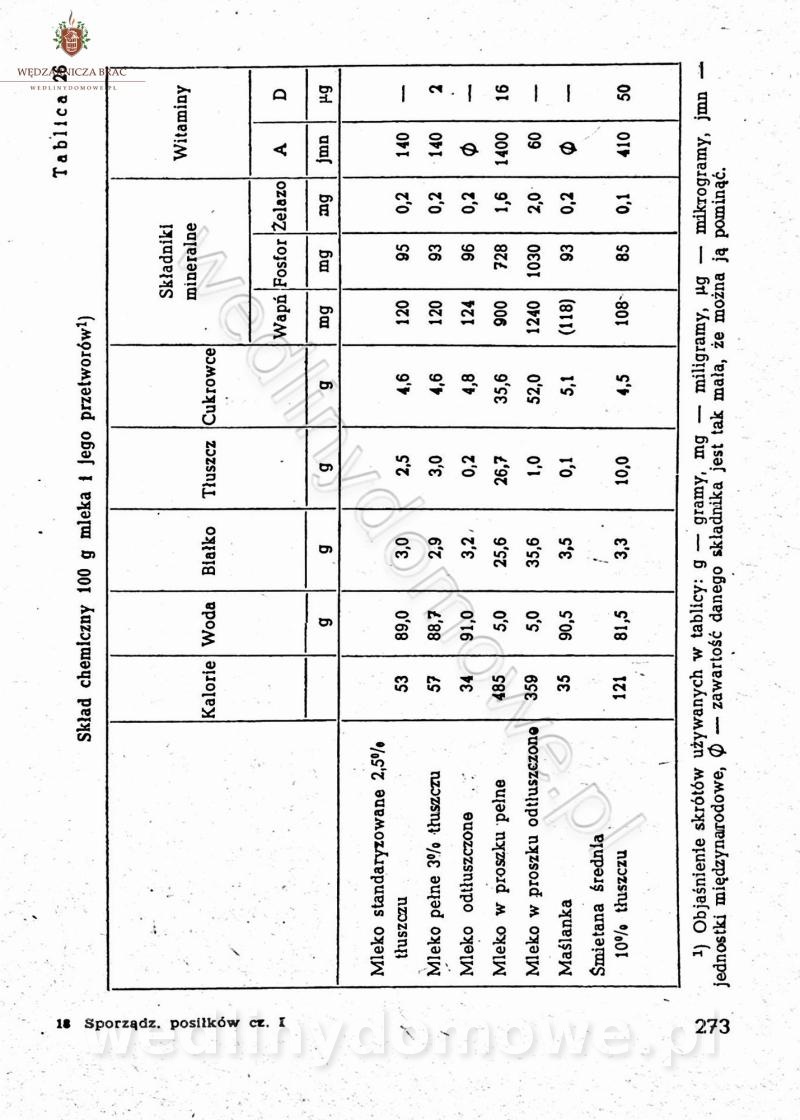
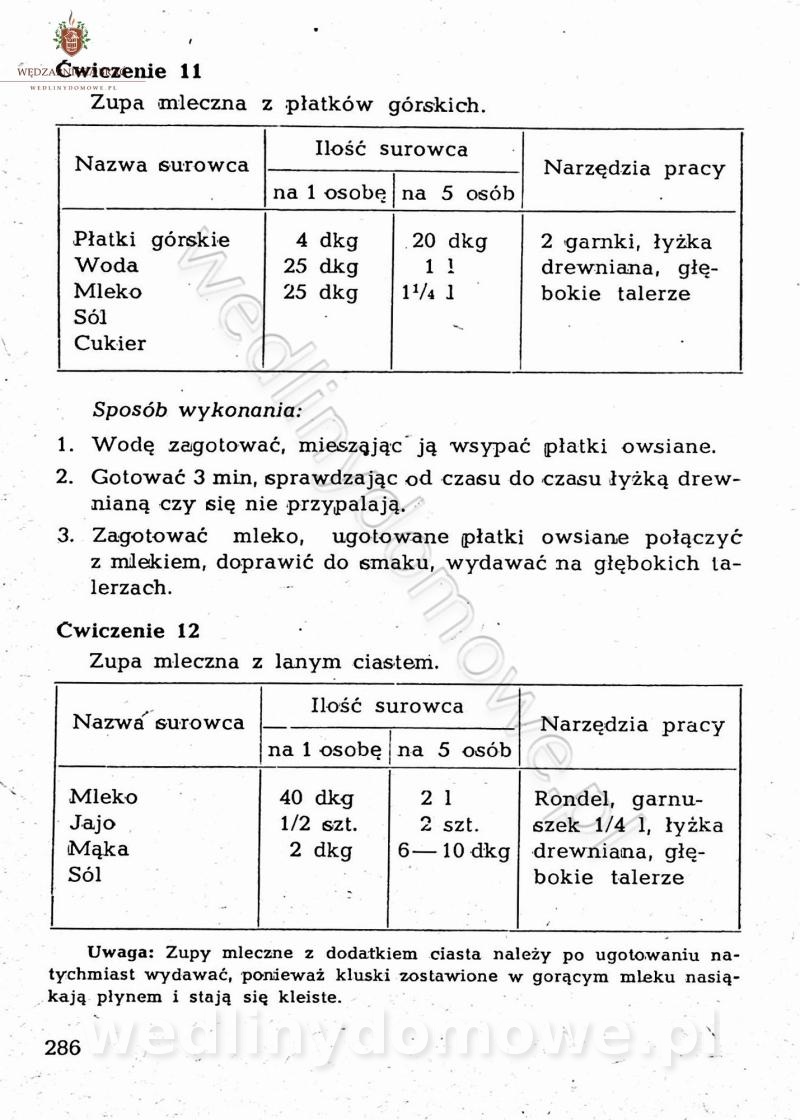
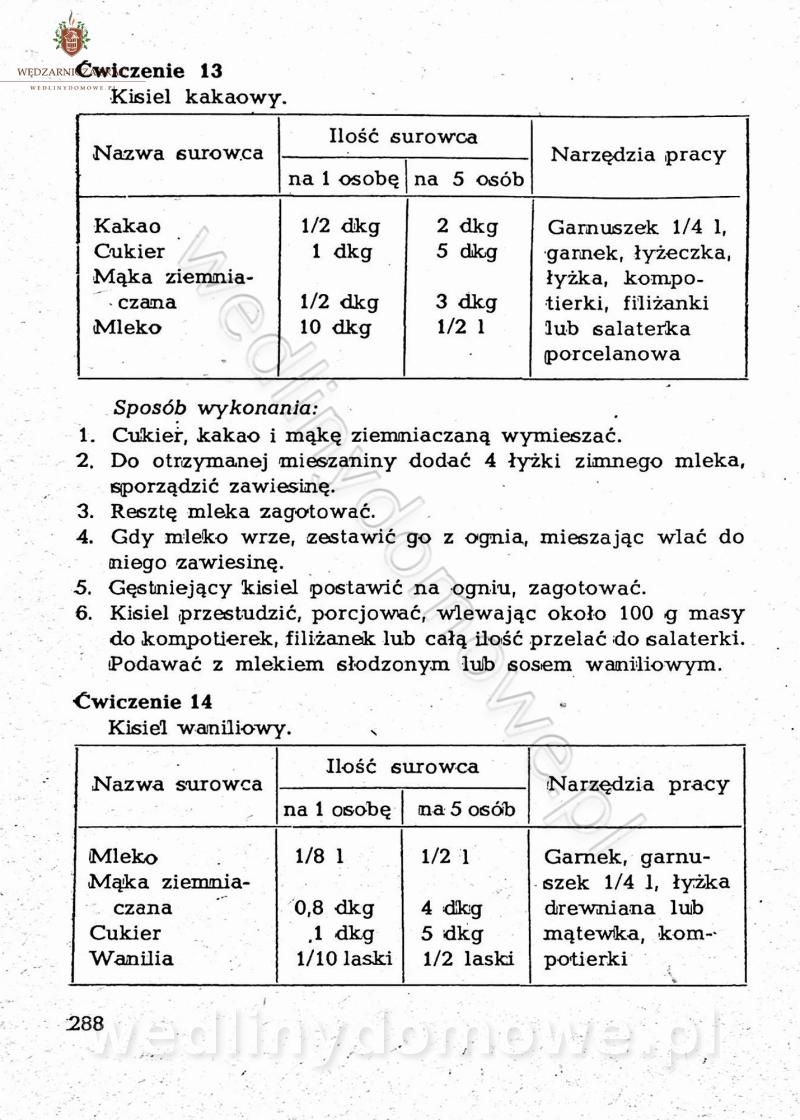
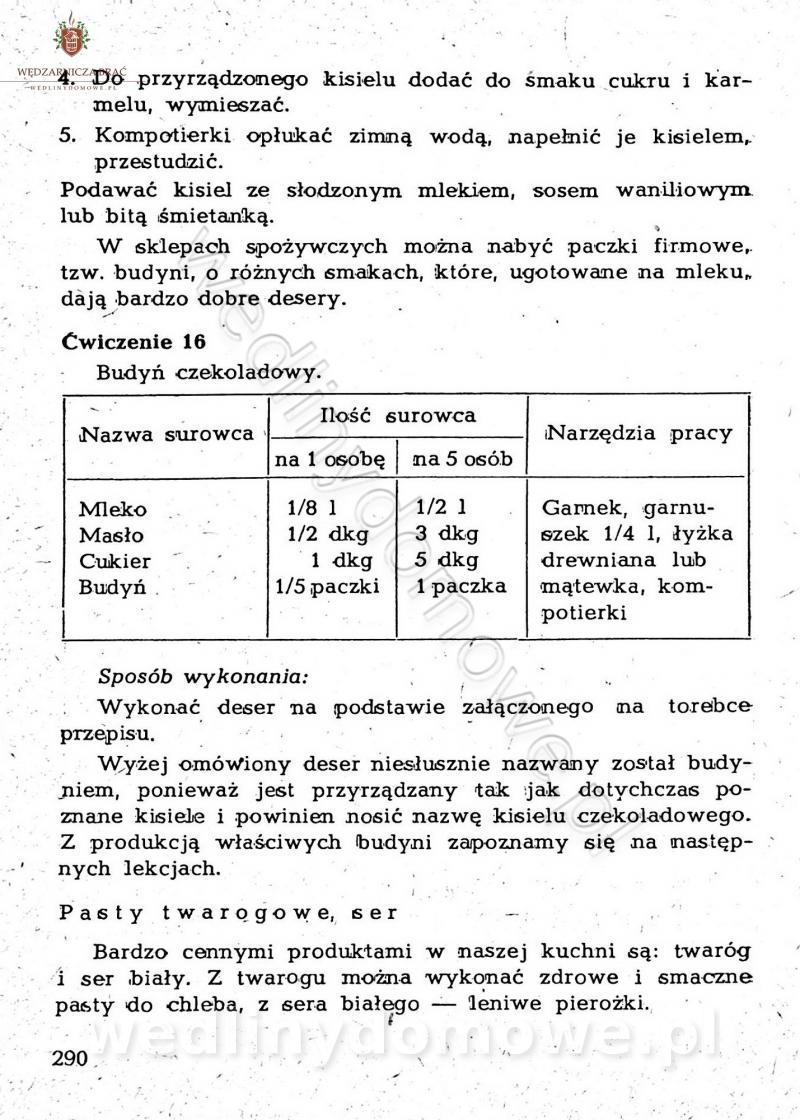
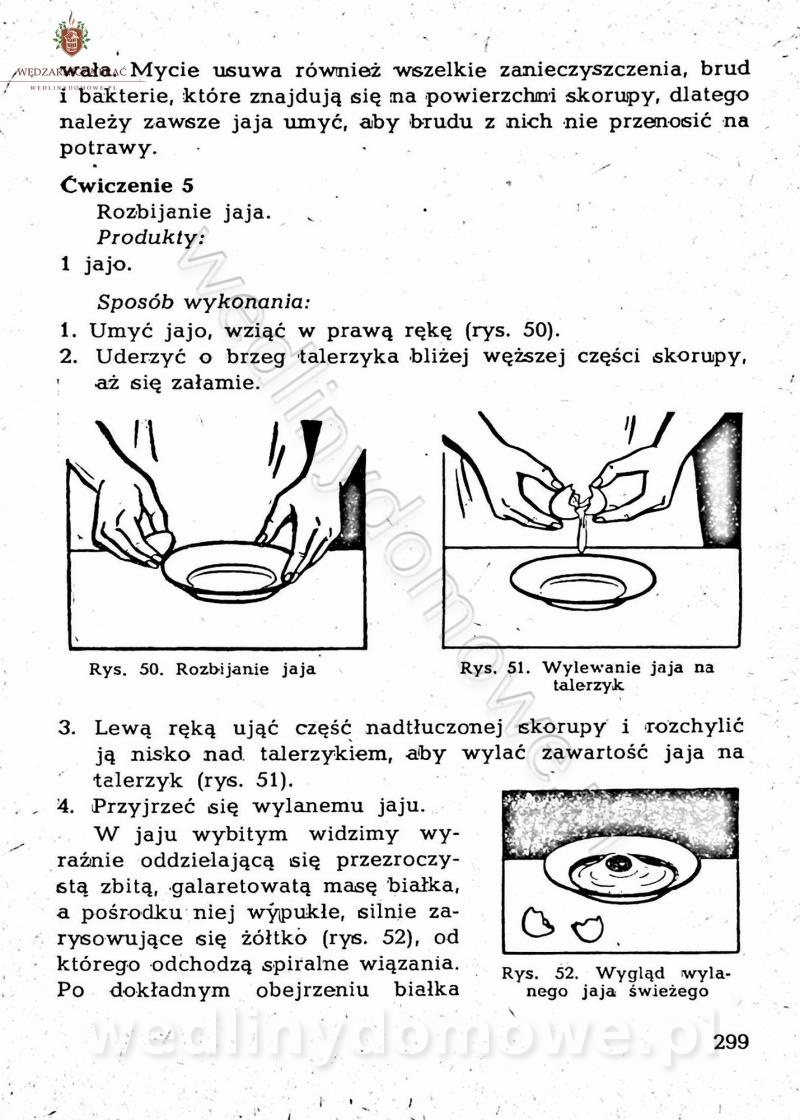
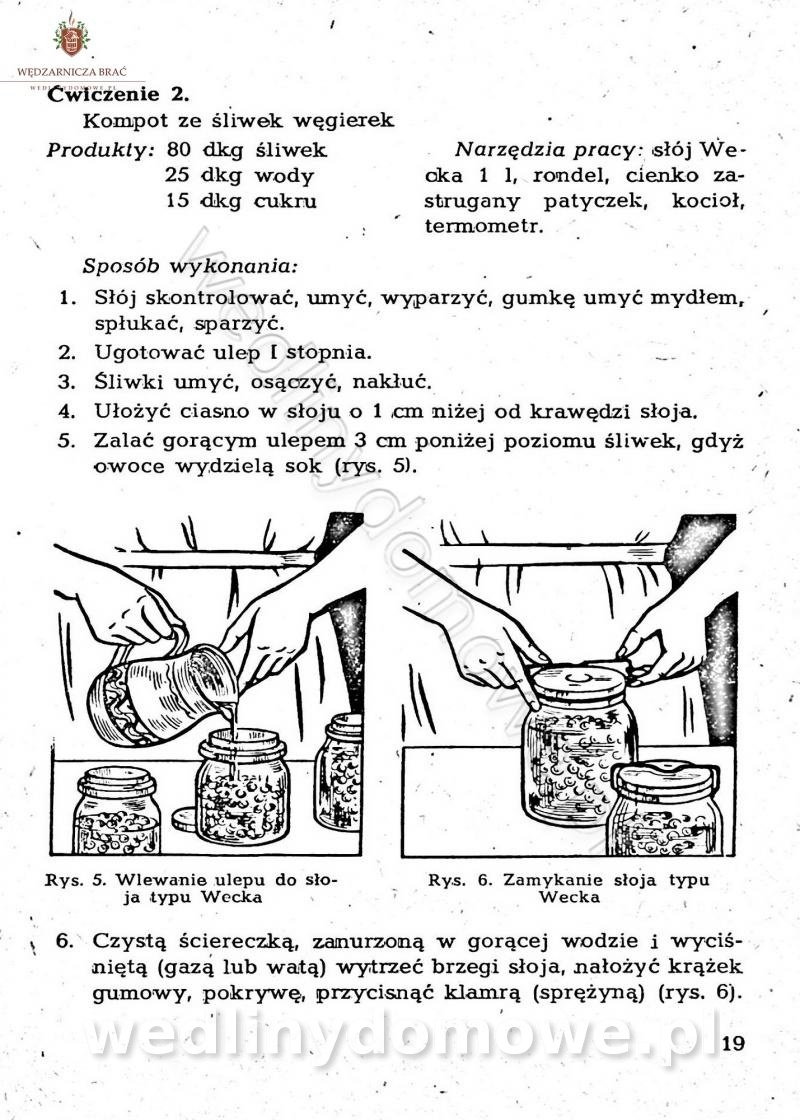
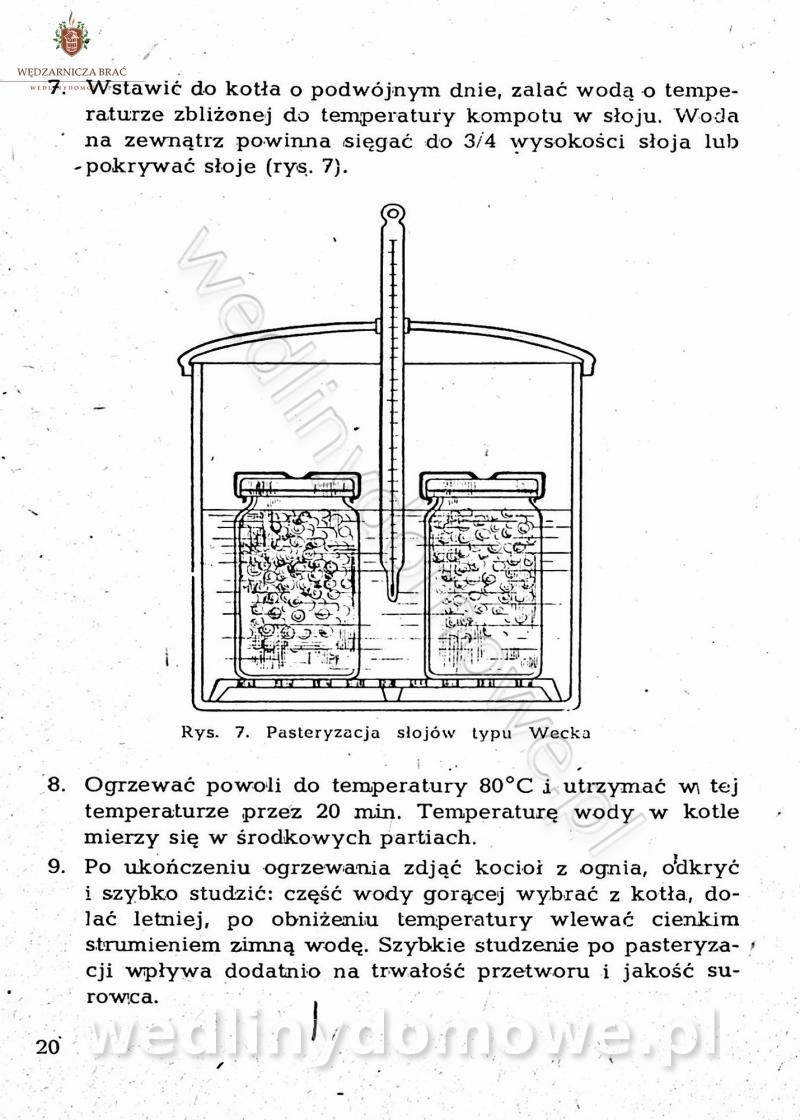
[Rok 1958][Tom 1] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie
Na potrzeby portalu wedlinydomowe.pl opracował Maxell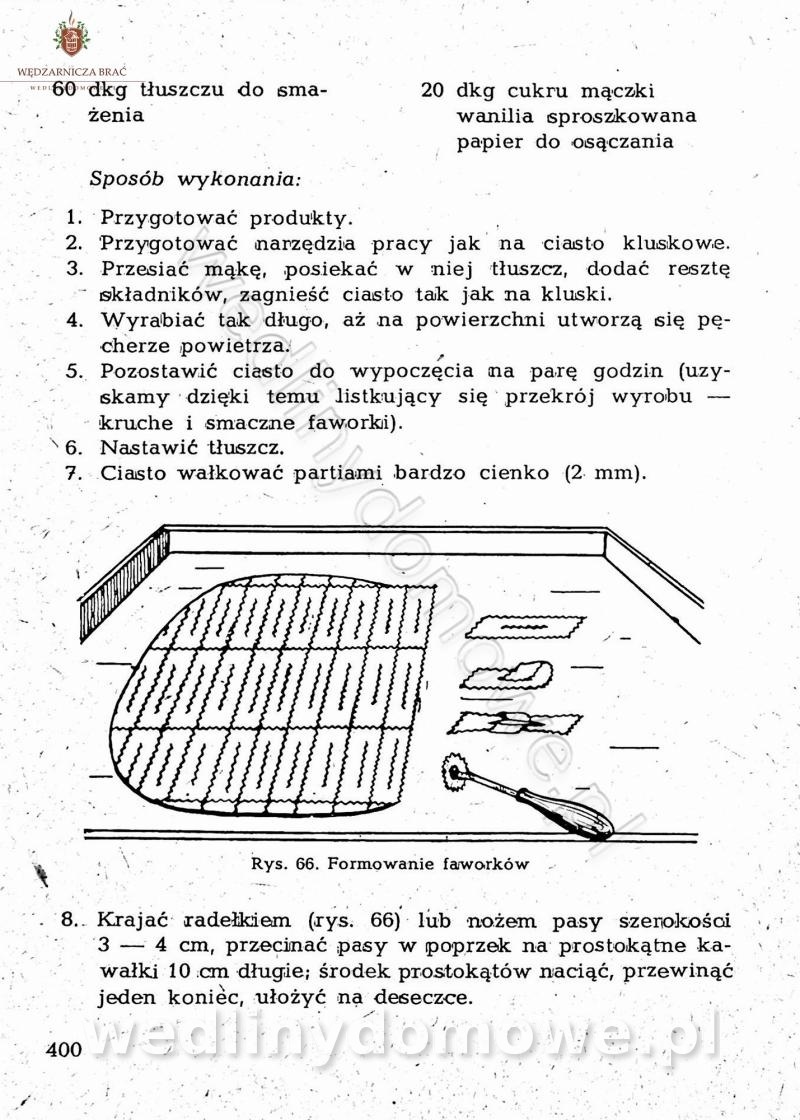





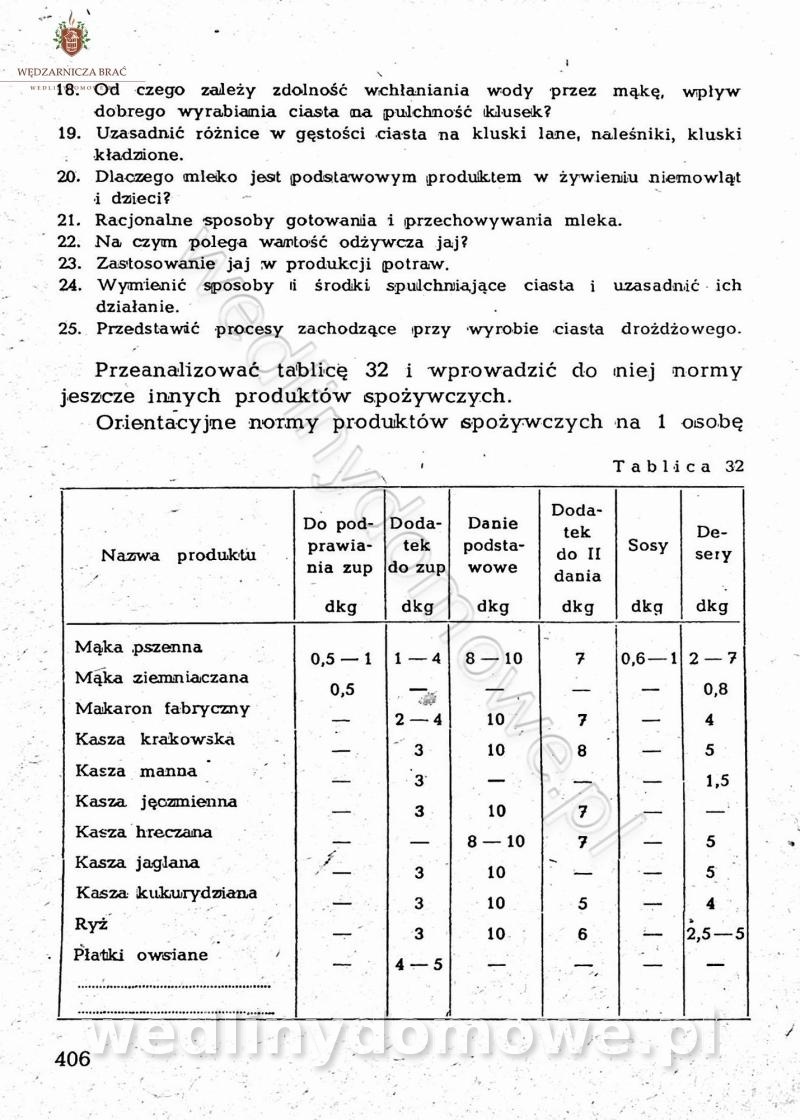

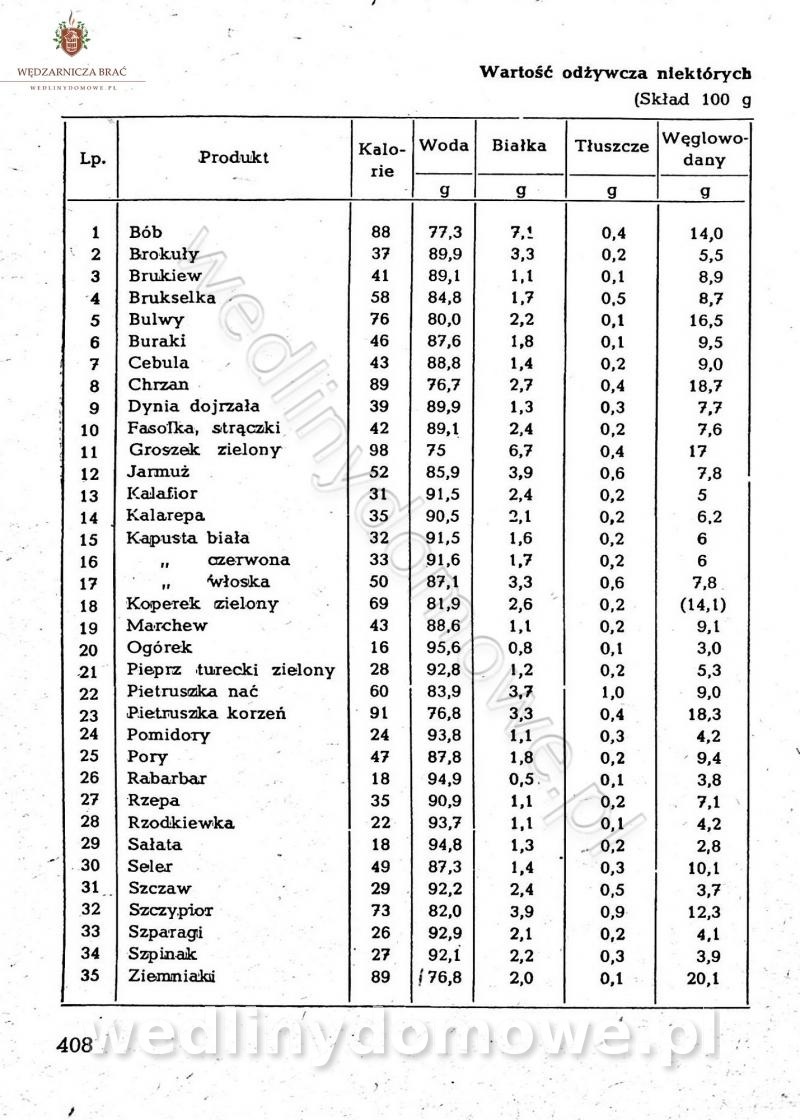
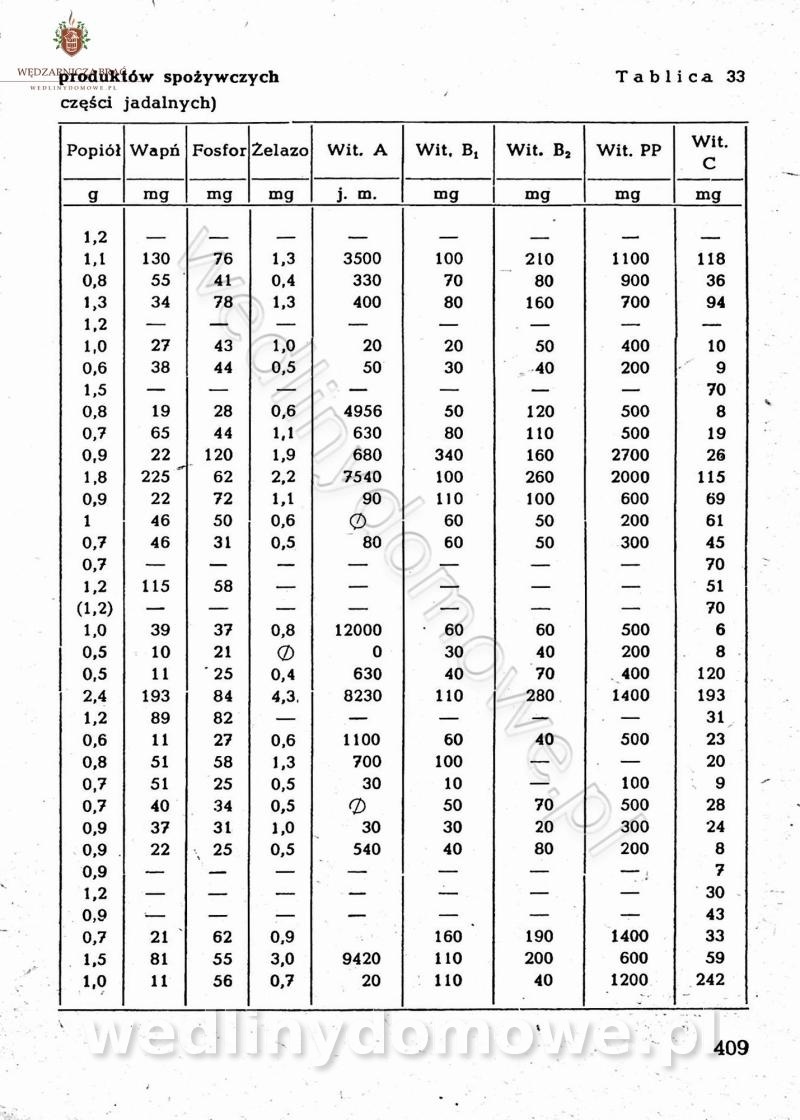
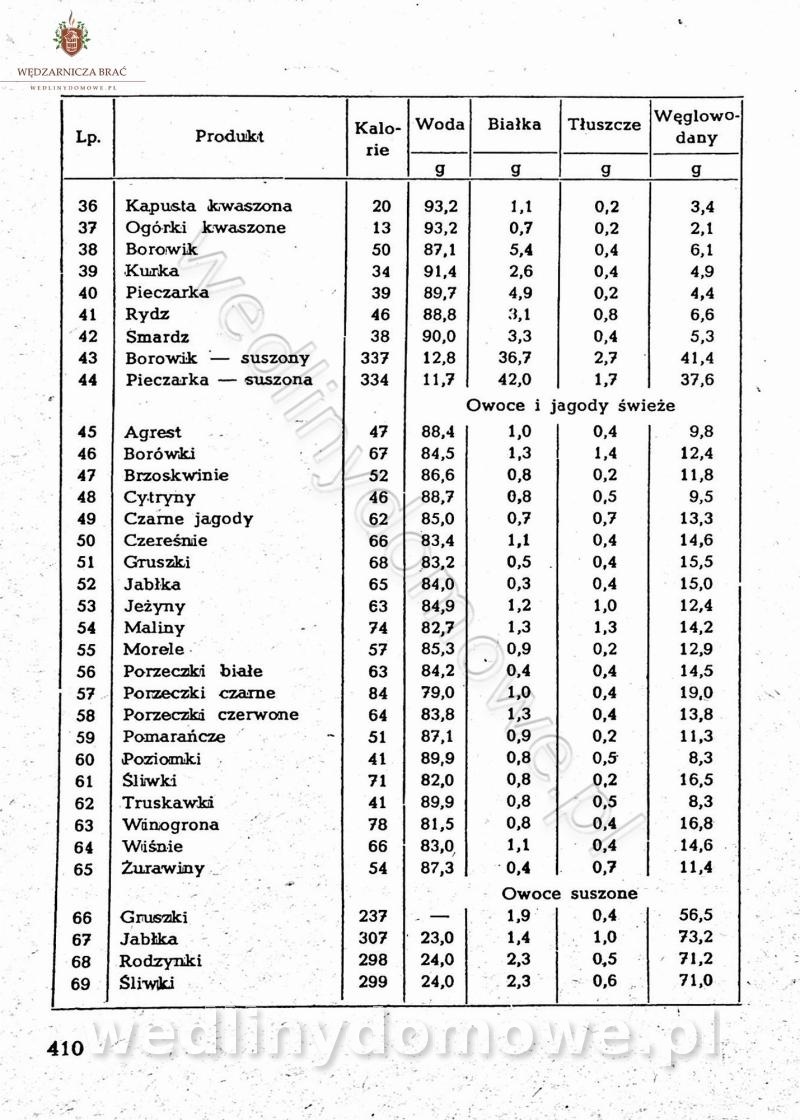
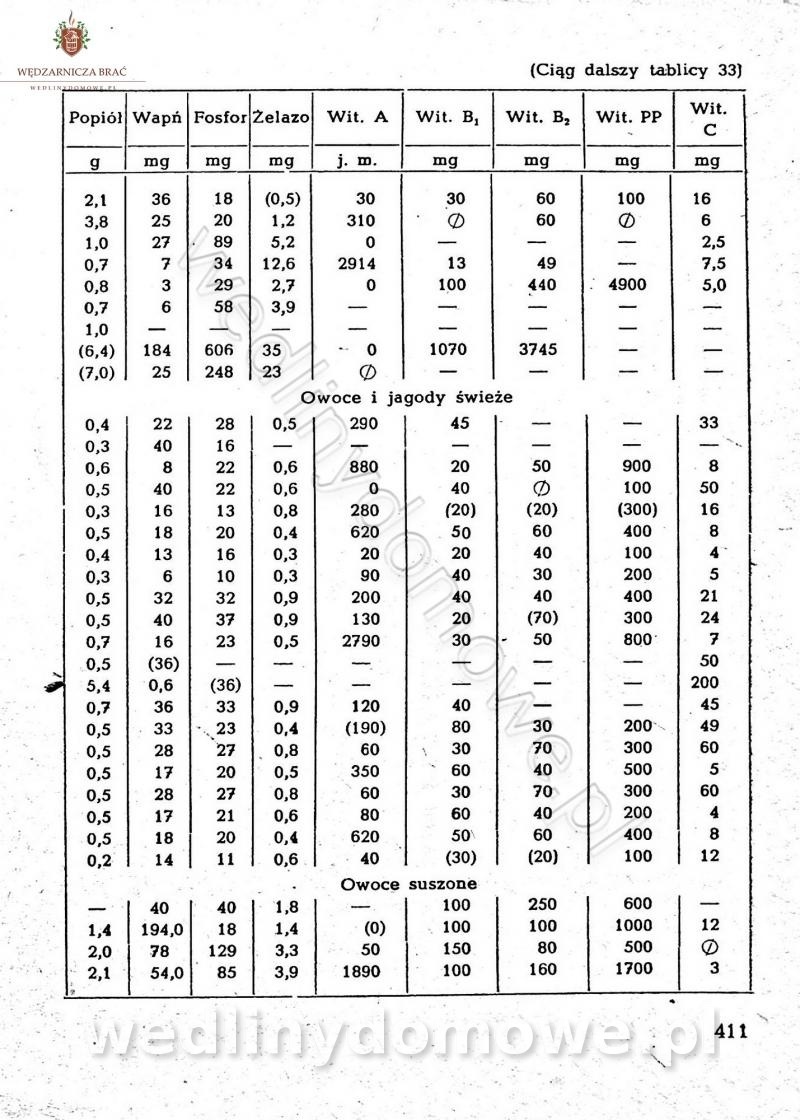
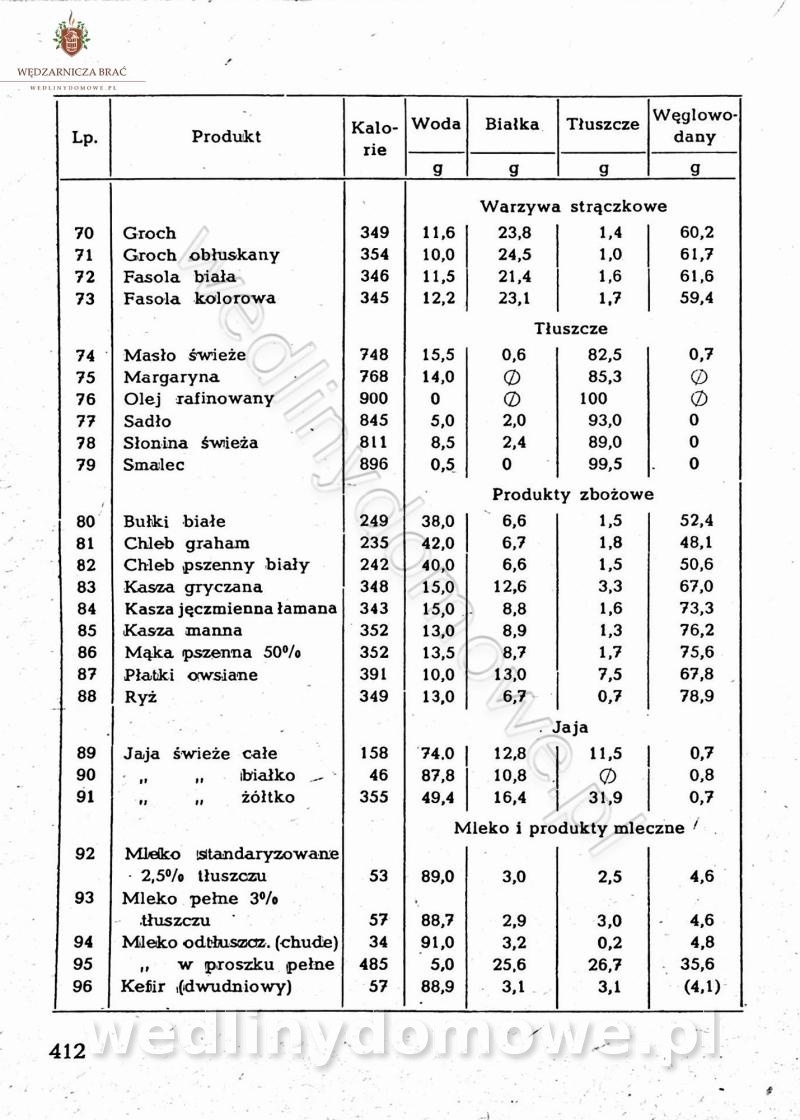
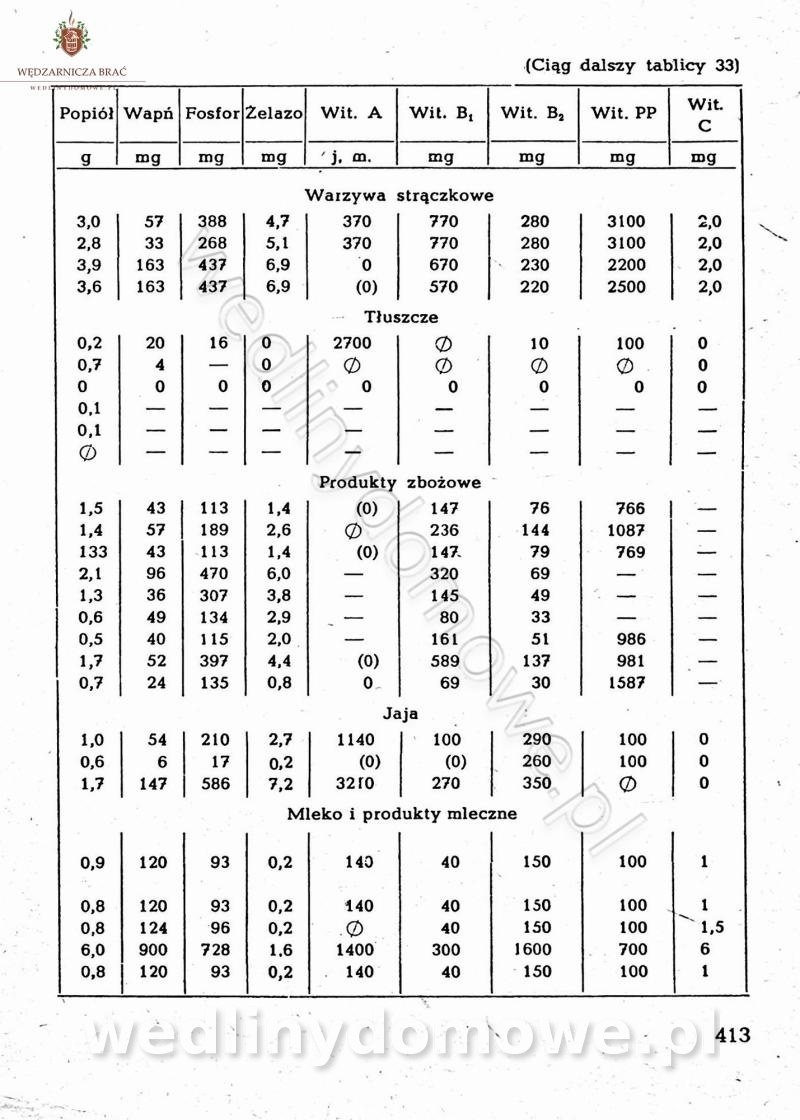
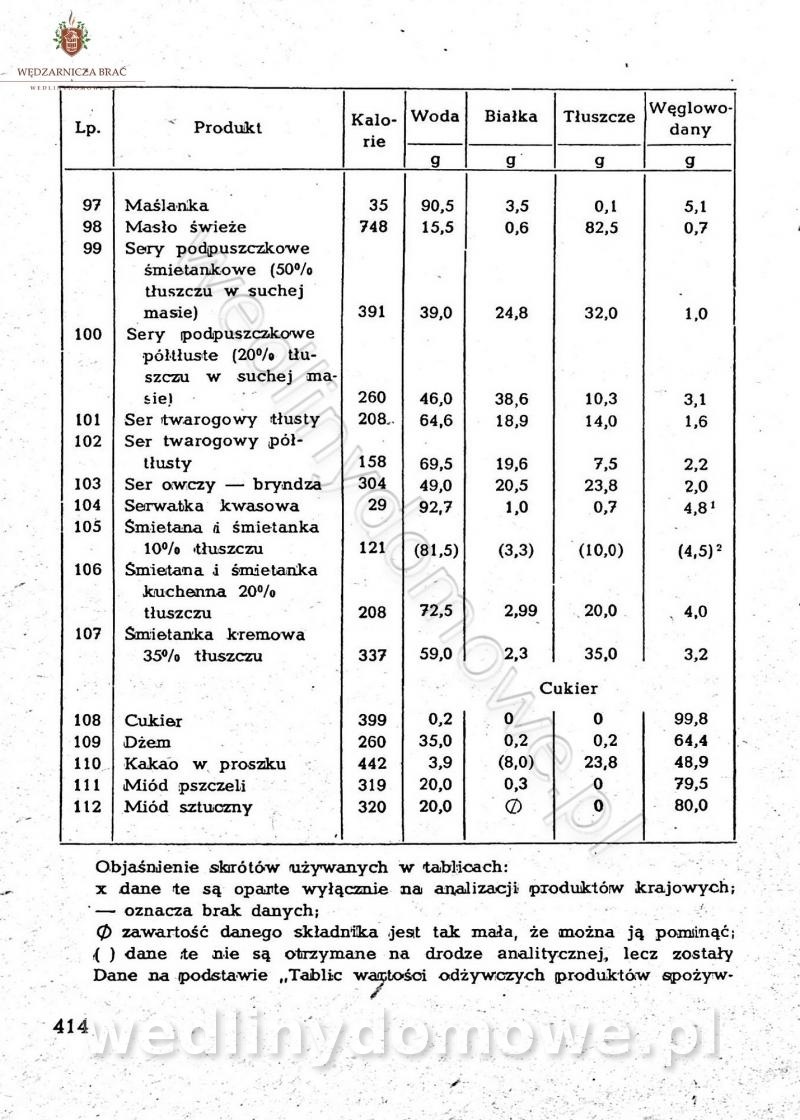
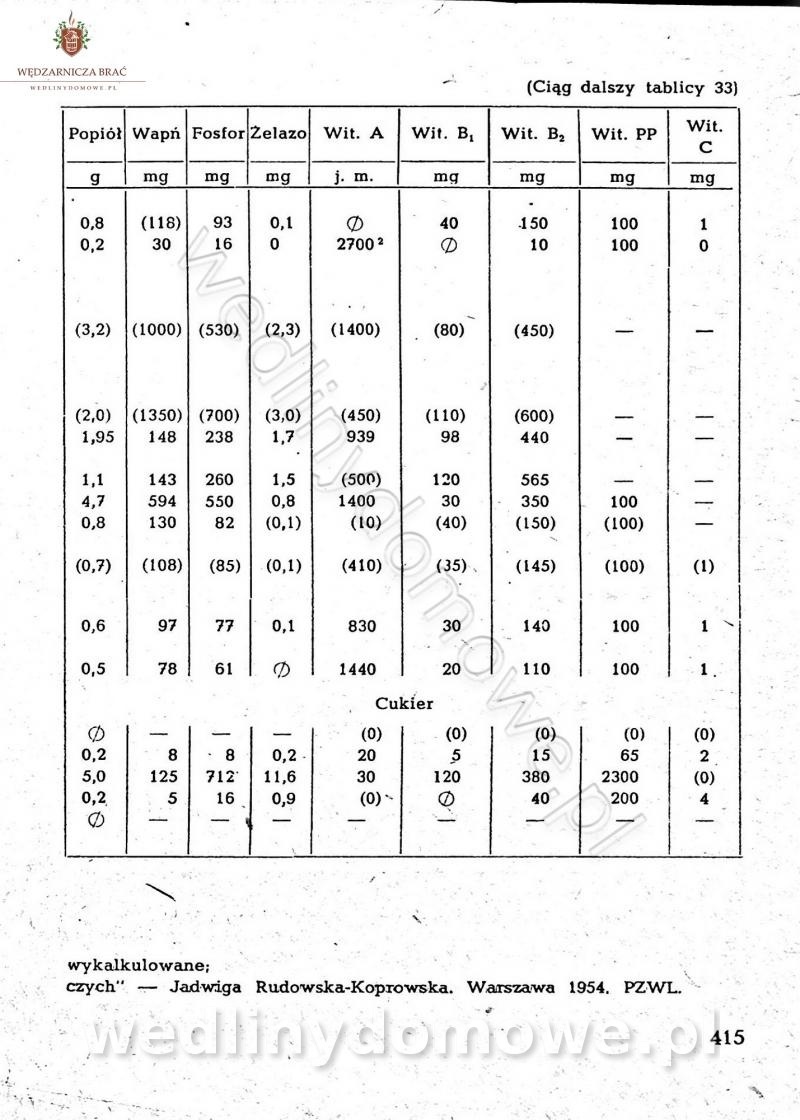







-
[Rok 1958][Tom 1] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie







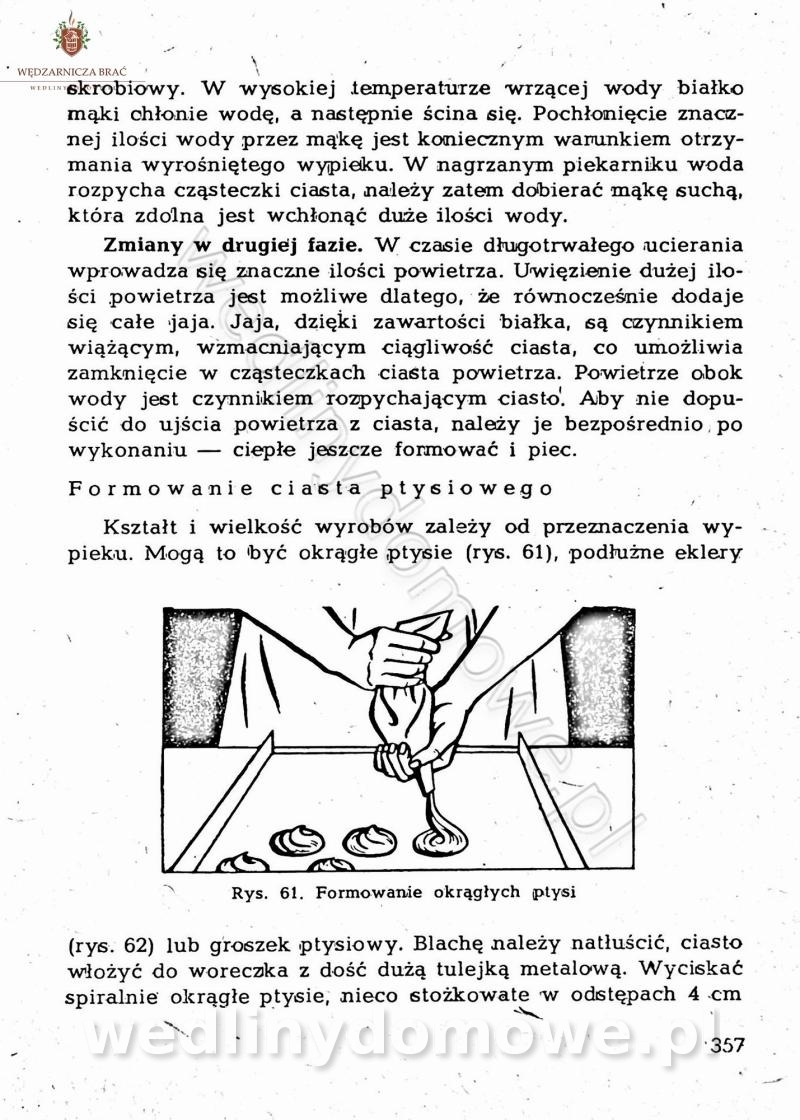
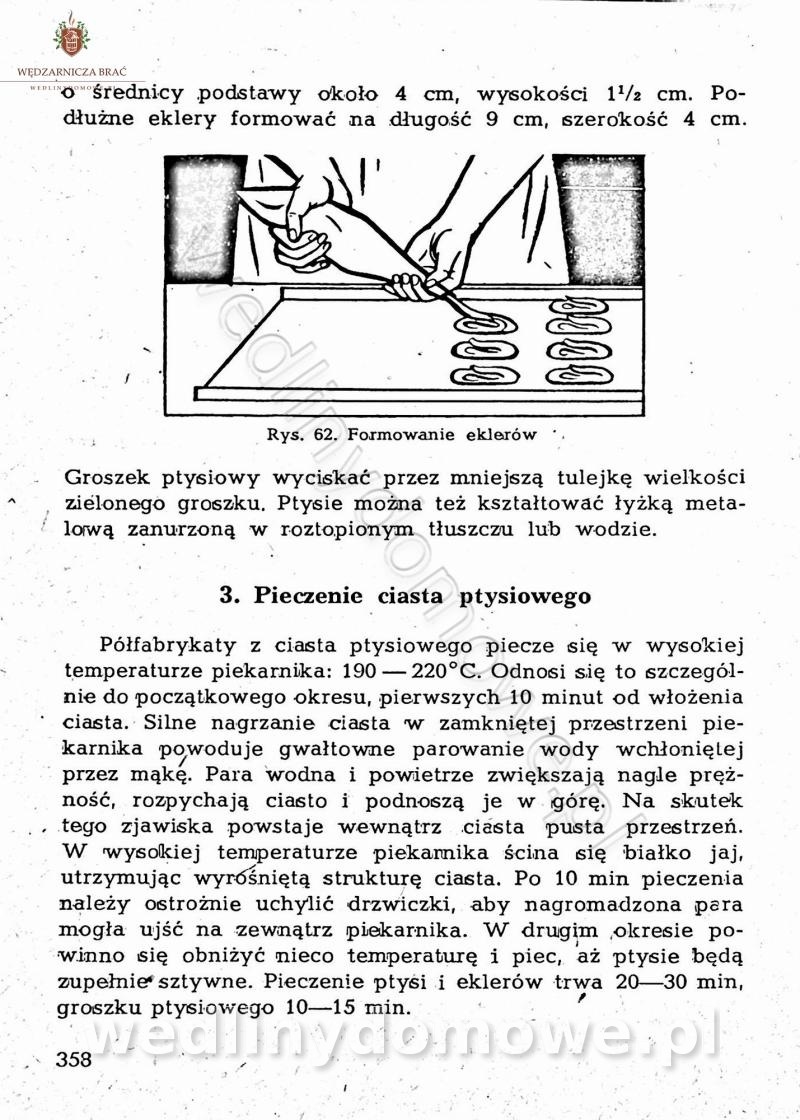






















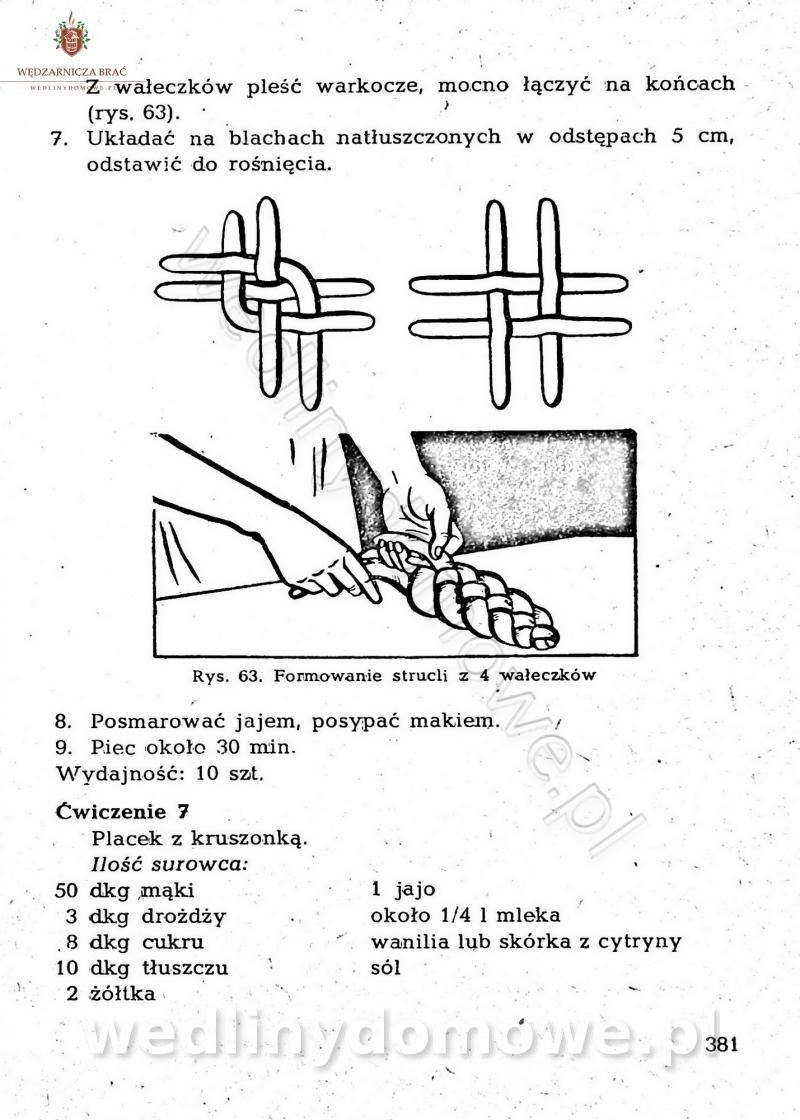


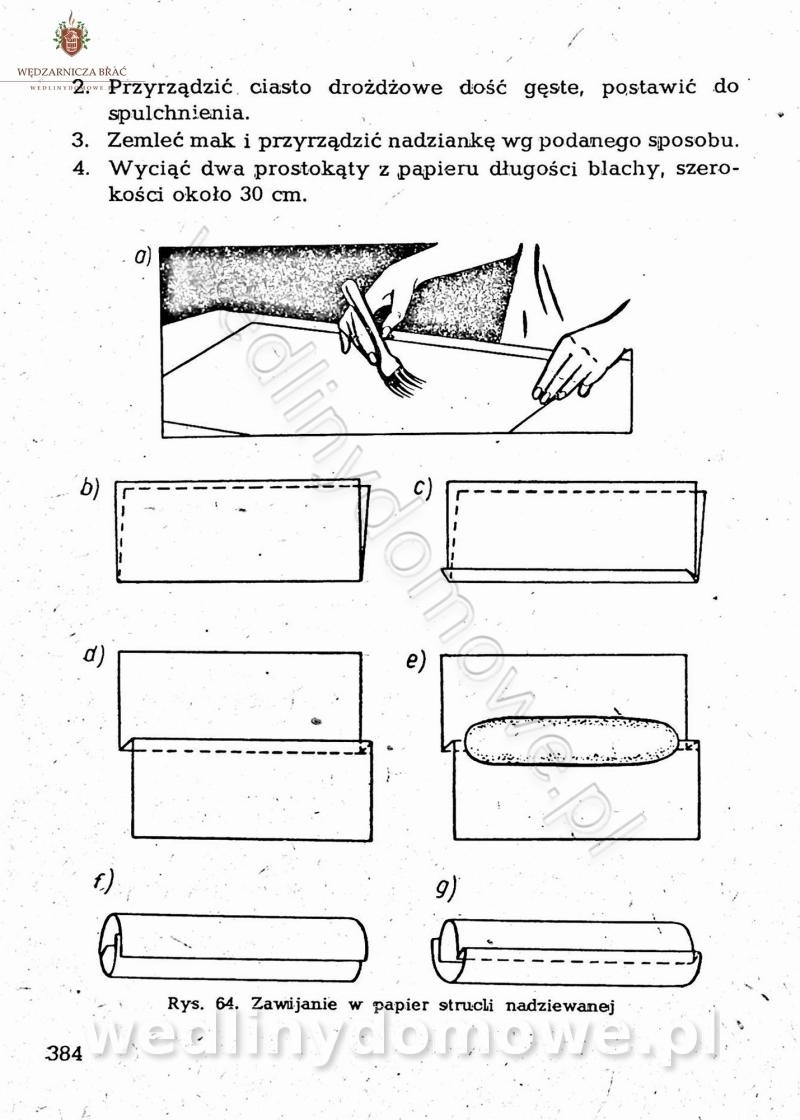















-
Szybkowar/sterylizator - Autoclav ALL AMERICAN model 925
Maxell odpowiedział(a) na EAnna temat w Sterylizatory
Składa się z trzech elementów o różnej masie. -
[Rok 1958][Tom 1] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie





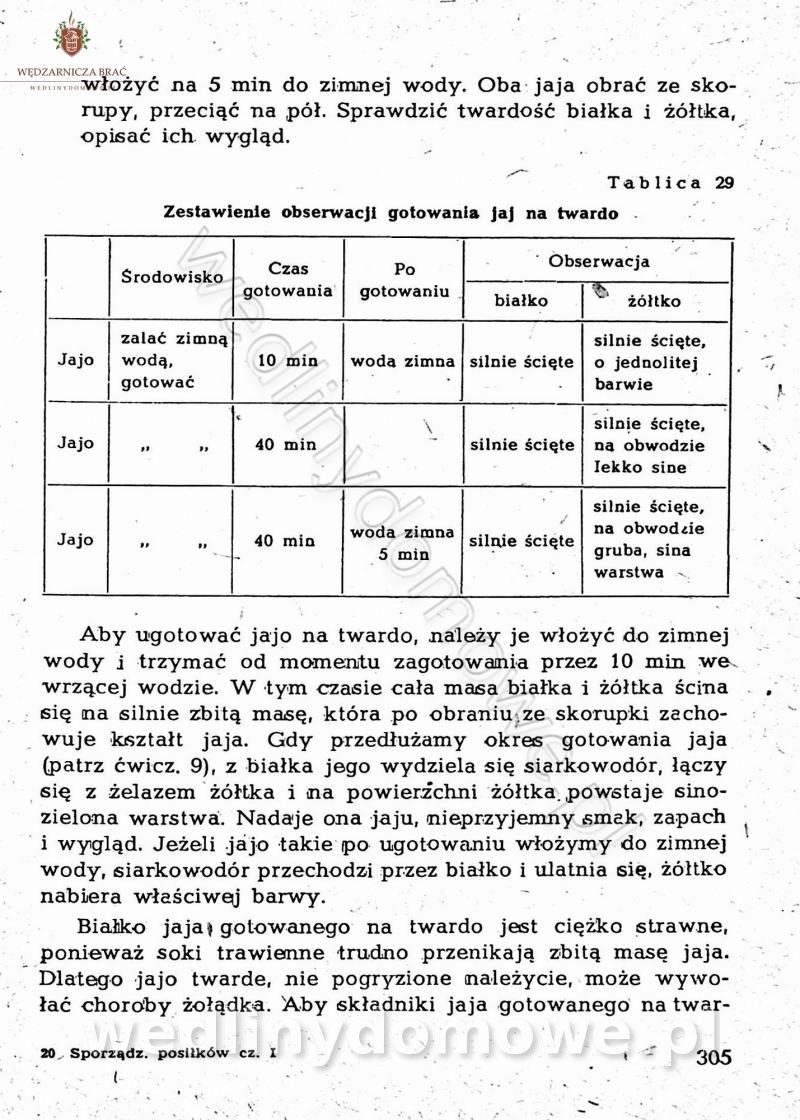







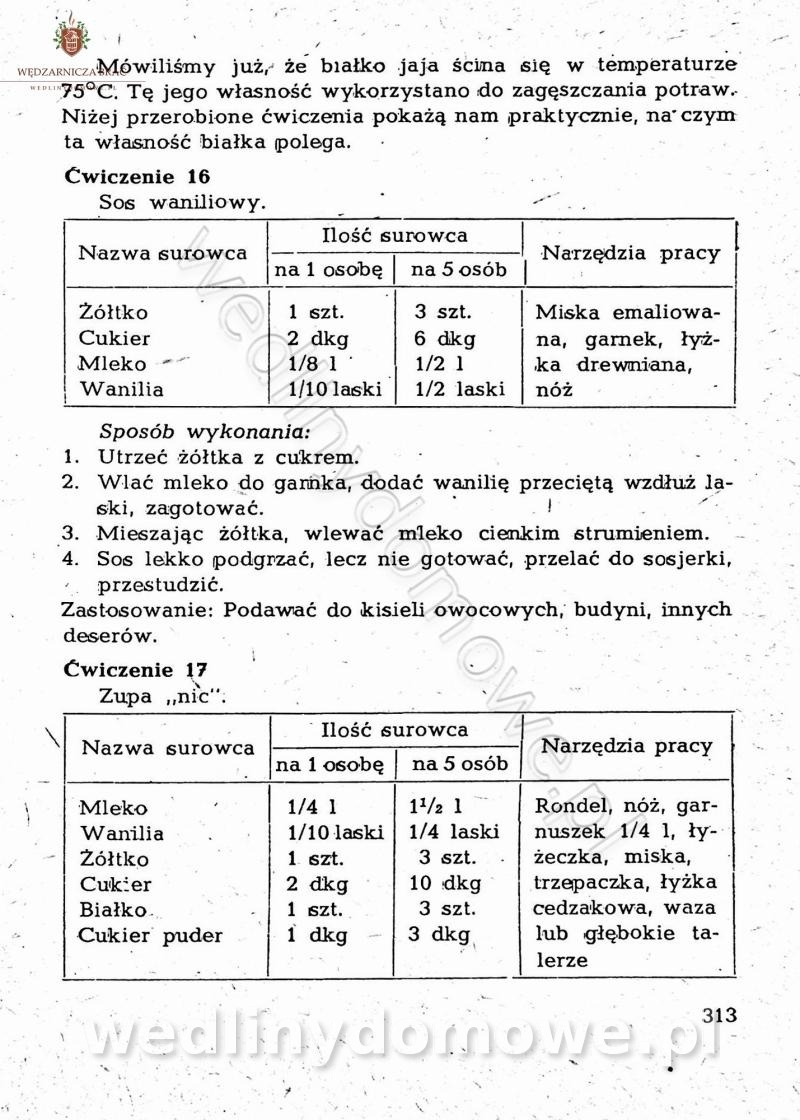





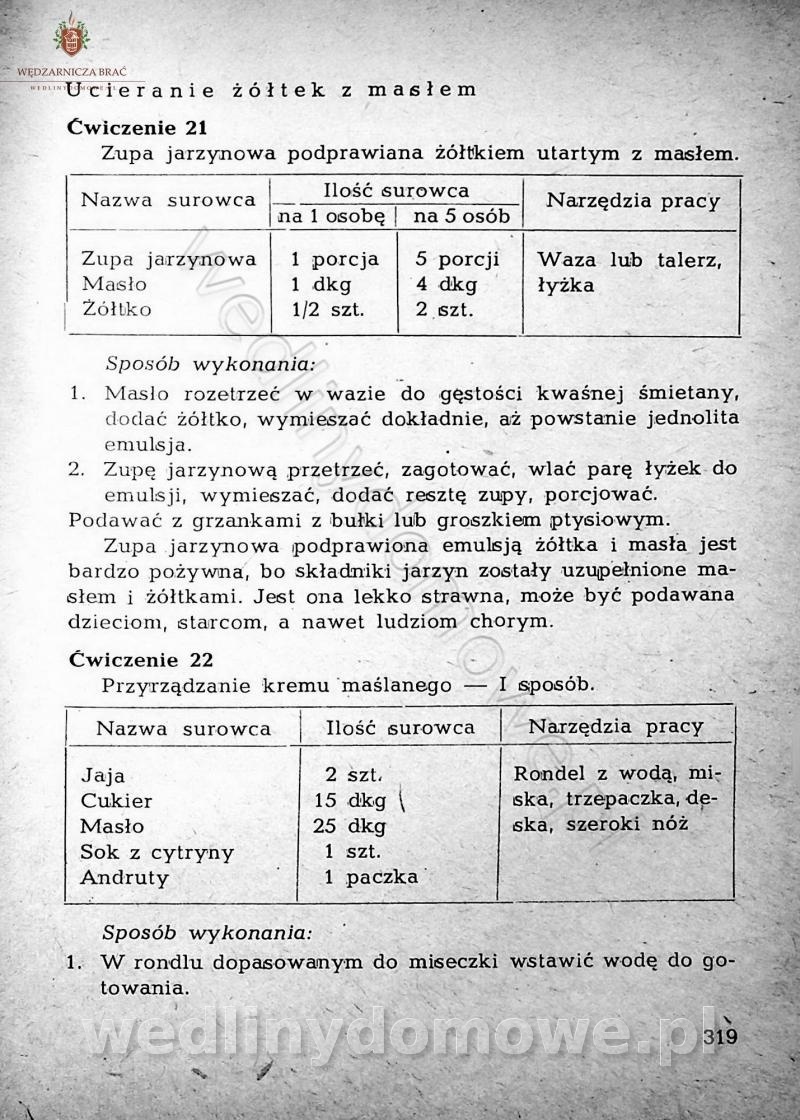



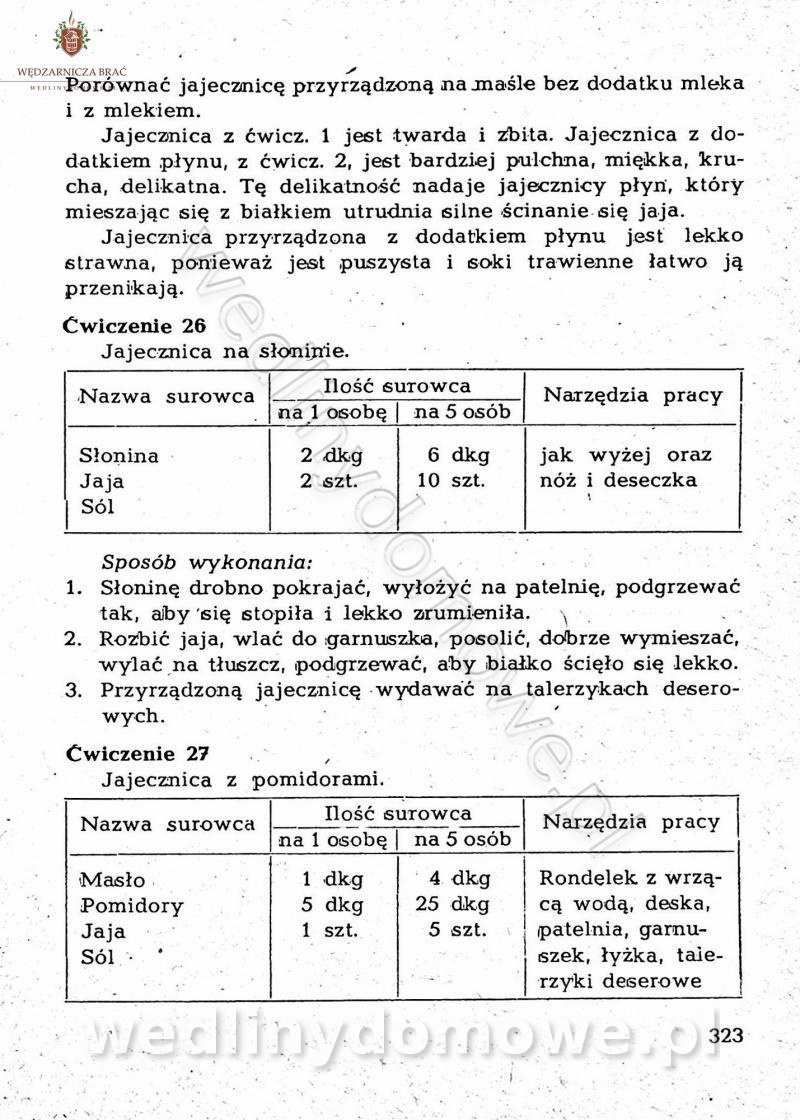
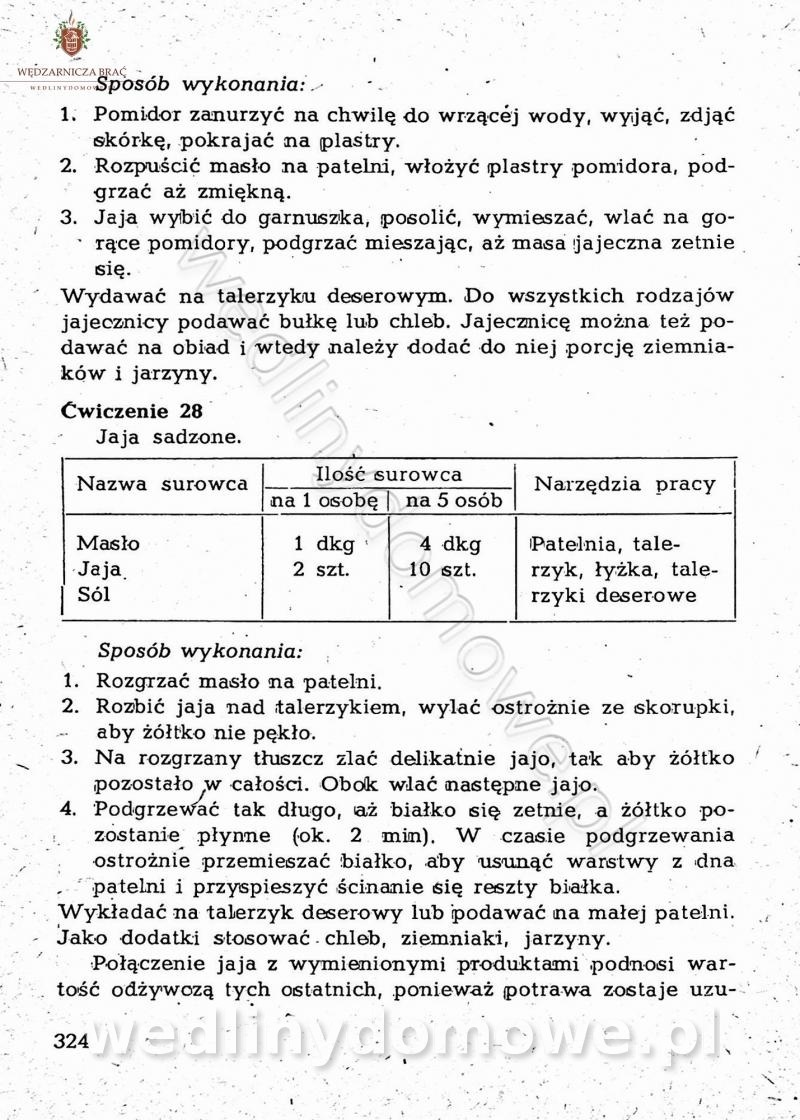









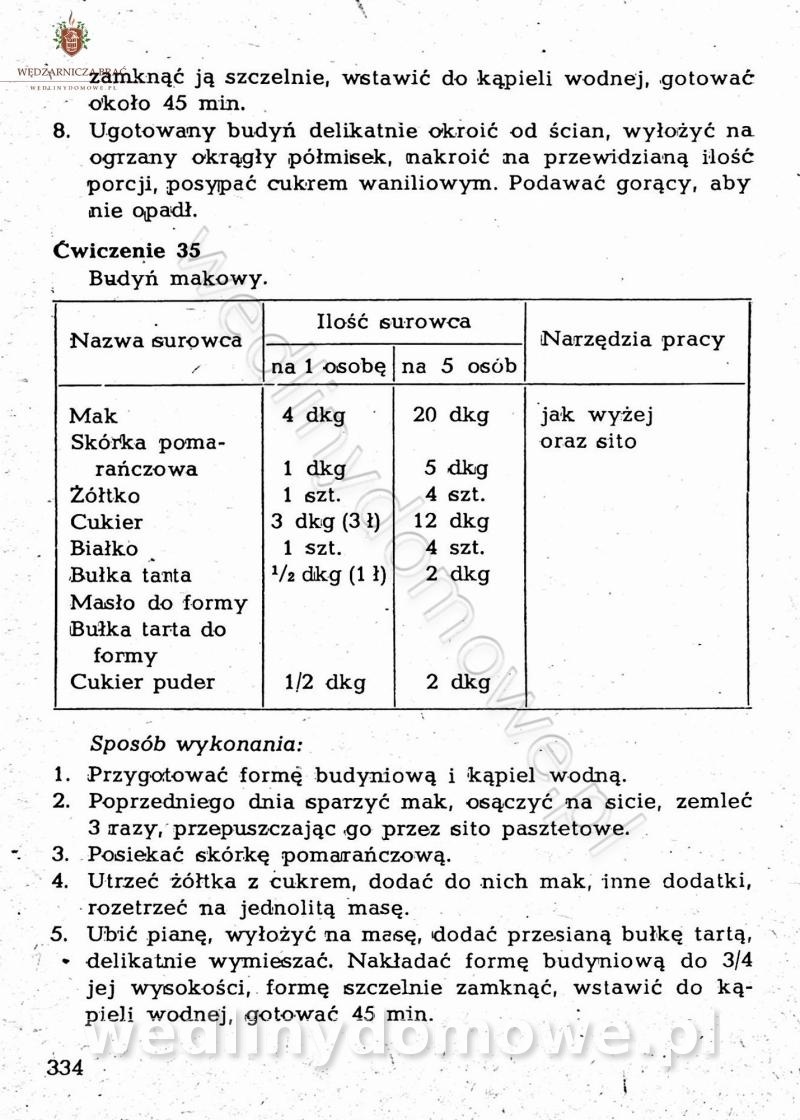















-
[Rok 1958][Tom 1] Sporządzanie posiłków.
Maxell odpowiedział(a) na Maxell temat w Książki kucharskie